Строение ротовой полости и отсутствие челюстных зубов и слюны исключают возможность существенной переработки пищи во рту. Заглоченную пищу карп измельчает с помощью глоточных зубов, которые способны перетирать твердые частицы до очень мелких размеров при многократно повторяющихся захватах и выплевывании.
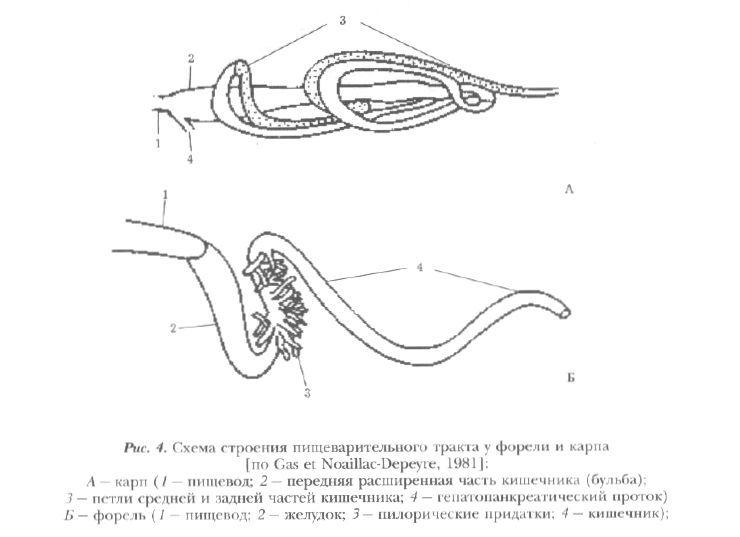
Из глотки пища поступает в короткий пищевод, а затем — в кишечник. Кишечник у карпа представляет длинную, в передней части заметно расширенную, а затем постепенно суживающуюся трубку, которая образует 8 петель. Длина кишечника превышает длину тела в 2-3 раза. Слизистая оболочка образует на его поверхности ячеистую структуру. Высота и число складок в переднем отделе приблизительно в 2 раза больше, чем в последующих. Складки увеличивают всасывающую поверхность кишечника в 9-12 раз [Ланге, 1948].
При наполнении пищей передняя часть кишечника способна сильно растягиваться, резко увеличиваясь в объеме. В связи с отсутствием сфинктеров пища непрерывно поступает в последующие его участки. Скорость продвижения пищи по кишечнику находится в прямой зависимости от температуры воды и в определенной связи с концентрацией кислорода и массой рыб. В качестве иллюстрации приводим диаграмму (см. рис. 1 Приложения), составленную Н.В. Рекубратским на базе экспериментальных данных [Рекубратский и др., 1985].
Проглоченная пища быстро вступает в контакт с желчью и пищеварительными соками, превращаясь в химус. Процесс переваривания происходит в среде с высоким и относительно постоянным содержанием воды (79-87%) при максимуме в переднем отделе. Реакция среды колеблется в пределах pH 6,1-7,6. Биохимический состав химуса, зависящий от уровня питательных веществ в корме, имеет определенные тенденции изменений: снижение к анусу относительного содержания в сухом веществе азотистых соединений и липидов и возрастание углеводов и минеральных элементов. При низком уровне в корме протеина (< 15%), липидов (< 2%), а также минеральных элементов их процент в химусе переднего отдела, как правило, существенно выше, чем в корме.
Главный источник пищеварительных ферментов — поджелудочная железа. Она не является компактным органом, а вкраплена в ткань печени (особенно со стороны, которая обращена к петлям кишечника) и встречается всюду, где имеются ветви воротной вены (на поверхности кишечника, в его мышечной оболочке, селезенке, печени и внутри ее). Она сопровождает кишечник вплоть до анального отверстия. Главный проток впадает вместе с общим желчным протоком сразу же за пищеводом недалеко от передней границы кишки. Мелкие протоки изливают свое содержимое по всей длине кишечника [Веригина, Жолдасова, 1982]. Отделение поджелудочного и кишечного соков происходит непрерывно вне зависимости от присутствия или отсутствия пищи. С началом питания секреция усиливается [Пегель, 1950].
Поджелудочная железа выделяет три группы высокоактивных ферментов, которые гидролизуют (расщепляют с присоединением воды) белки, жиры, углеводы: протеазы (трипсин, химотрипсин, различные пептидазы), амилазу, липазу.
Большую роль в пищеварении и в обмене веществ в целом играет печень. Она представляет собой объемистый многолопастной орган, в который погружен кишечник. Желчный пузырь помещается на внутренней стороне главной дольки печени и полностью покрыт ею. Главный секрет печени — желчь. Выделяясь непрерывно, она при ограничении или отсутствии пищи концентрируется в желчном пузыре.
Желчь, облегчая всасывание жиров, одновременно стимулирует секреторную функцию поджелудочной железы, а также тонус и моторику переднего отдела кишечника и препятствует развитию гнилостной микрофлоры.
Другим важным источником пищеварительных ферментов служит слизистая оболочка кишечника. Ее железы выделяют слизь, содержащую мукополисахариды, которая обволакивает пищу и облегчает ее продвижение по кишечнику.
На всем протяжении кишечника его поверхность выстлана высоким однослойным цилиндрическим каемчатым эпителием, клетки которого называются энтероцитами или эпителоцитами. Они продуцируют ферменты, которые выделяются в полость или попадают туда со слущивающимся эпителием. Обновление энтероцитов передней половины кишечника, осуществляющей наиболее активное пищеварение, у карповых рыб происходит за неделю, задней — за 5-7 недель. При этом весной и летом скорость обновления выше, чем зимой. Энтероциты кишечника карпа имеют строение, сходное с их строением у высших позвоночных [Noaillac-Depeyre, Gas, 1973].
С помощью панкреатических и кишечных ферментов осуществляется полостное пищеварение. Сущность его заключается в том, что в полости кишки происходит ферментативное расщепление пищи, и когда его продукты достигают поверхности слизистой оболочки, начинается всасывание нутриентов (питательных веществ, расщепленных до мономеров, т. е. аминокислот и жирных кислот, моносахаридов, витаминов и т. д.).
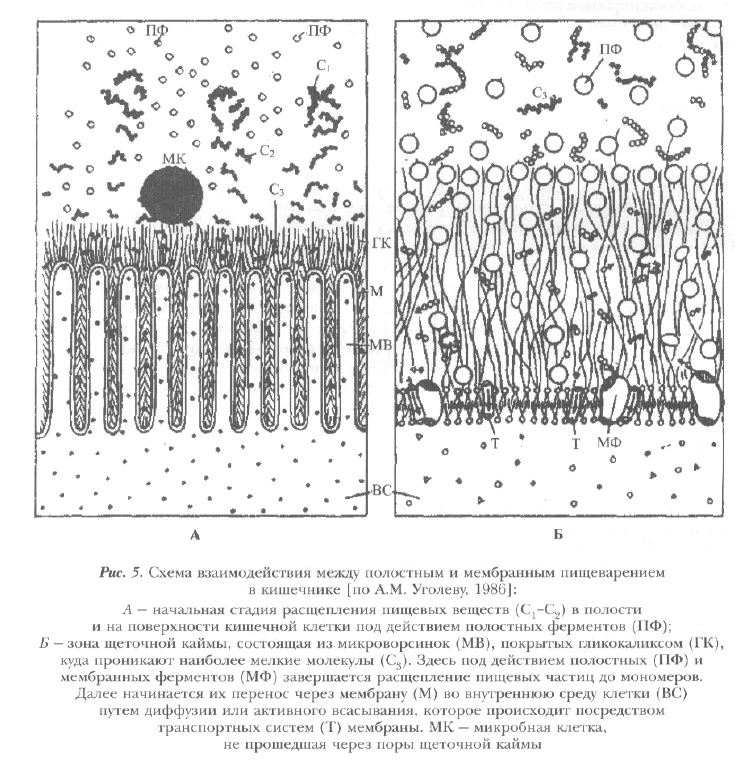
Часть ферментов, секретируемых кишечными клетками, и ферментов панкреатического происхождения адсорбируется на выростах внешней поверхности энтероцитов — "щеточной кайме" или "микроворсинках", покрытых гликокаликсом — дополнительным предмембранным слоем из тонких извитых нитей, которые заполняют и поры между микроворсинками (рис. 5).
Микроворсинки являются структурной основой другого мощного механизма пищеварения, свойственного всем позвоночным. Он был открыт А.М. Уголевым в конце 50-х годов и имеет несколько названий, отражающих его сущность: "пристеночное", "контактное", "мембранное" [Уголев, 1960].
Этот тип пищеварения играет промежуточную роль в механизме усвоения пищи. Он осуществляется с помощью ферментов, адсорбированных на микроворсинках, т.е. на границе внеклеточной и внутриклеточной сред. Микроворсинки образуют на поверхности мембран энтероцитов ультрапористость, что в десятки раз увеличивает поверхность контакта пищи со всасывающими клетками. Таким образом создаются структурные предпосылки для быстрого расщепления и одновременно всасывания биополимеров в этой зоне. В результате процессы гидролиза и транспорта становятся сопряженными в пространстве и времени.
Между полостным и мембранным пищеварением существуют определенные взаимоотношения. В начальных отделах пищеварительного тракта (ротовой полости, глотке, пищеводе, желудке) пища, представляющая собой животные и растительные ткани, подвергается первичной механической и химической обработке. Ферменты, находящиеся в полости, через поврежденные клеточные оболочки тканей пищи быстро расщепляют клеточное содержимое на крупные и мелкие фрагменты. Продукты распада диффундируют за пределы клеток пищи и, передвигаясь с помощью перистальтики, достигают поверхности слизистой оболочки. Здесь в зоне пристеночного пищеварения измельченные частицы подвергаются окончательному гидролизу с последующим всасыванием в зоне микроворсинок. Крупные частицы (более 100-200 А) отбрасываются в полость кишечника и вновь подвергаются воздействию полостных ферментов (см. рис. 5).
Перенос продуктов ферментативного расщепления через клеточную оболочку происходит как путем пассивного (т. е. диффузии), так и активного транспорта с помощью узкоспециализированных переносчиков (массой 10-70 тыс. Да), связывающих и перемещающих мономеры через мембраны с затратой энергии АТФ.
Первые сведения о наличии у рыб мембранного пищеварения на примере взаимодействия гидролитической и транспортной функции углеводов в кишечнике карпа были опубликованы Ш.А. Берман в 1964 г. К настоящему времени в этом направлении накоплен большой экспериментальный материал. Активные работы ведутся в институте биологии внутренних вод АН России [Уголев, Кузьмина, 1993; Кузьмина, 1995 и др.]
Переваривание пищи. Ранее предполагалось, что ферментативное расщепление и всасывание питательных веществ в пищеварительном тракте рыб разделены в пространстве и времени [Карзинкин, 1932; Пегель, 1950; Краюхин, 1963]. Однако благодаря использованию метода инертных веществ, позволившего количественно охарактеризовать степень переваривания основных нутриентов на последовательных участках пищеварительного тракта рыб, было показано, что всасывание продуктов расщепления питательных веществ происходит на всем протяжении кишечника [Щербина, 1967, 1969, 1970, 1971, 1973, 1975, 1980, 1983-1984].
С позиций пристеночного пищеварения получил объяснение факт очень интенсивной резорбции нутриентов в передней расширенной части кишечника карпа, на которую приходится около 14% его длины. Здесь извлекается из пищи и всасывается в среднем около ¼ сухих веществ, доступных рыбам, в том числе около 25% белка, от 20 до 80% отдельных аминокислот, до 90% простых сахаров, большое количество липидов. Таким образом, передний расширенный отдел кишечника, помимо депонирующей, активно осуществляет гидролитическую и резорбтивную функции. Высокая интенсивность всасывания органических веществ сохраняется еще на протяжении ⅓ длины кишечника. Объем резорбции азотсодержащих веществ и аминокислот может быть несколько выше или ниже. Преимущественно здесь локализован максимум всасывания липидов. Возрастает интенсивность всасывания магния, кальция и других минералов. Из этого следует, что отсутствие у карпа желудка и связанного с ним пепсинового пищеварения не замедляет, как предполагалось ранее [Карзинкин, 1950; Краюхин, 1963], темпов переваривания белков, липидов, полисахаридов, а также всасывания аминокислот и моносахаридов. Протео- и амилолитические ферменты, работающие в слабокислой и слабощелочной среде, гидролизуют белки и углеводы с очень высокой скоростью, обеспечивающей их столь же интенсивную резорбцию. Далее она резко сокращается и активизируется резорбция минеральных веществ.
В конечных участках кишечника (-10% его длины) завершаются процессы всасывания расщепленных питательных веществ. Идет активное всасывание воды, и пищевой химус превращается в экскременты — остатки непереваренной пищи, отмершие клетки кишечного эпителия, эндогенные вещества и бактериальную массу. Они обволакиваются большим количеством слизи и удаляются из организма под действием перистальческих движений кишечника. В экскрементах содержится более 85% воды, в их сухом веществе находится 7-17% сырого протеина (нередко и более), 50% и более углеводов, 15% и более минеральных веществ. Выделяются экскременты через анальное отверстие практически непрерывно.