
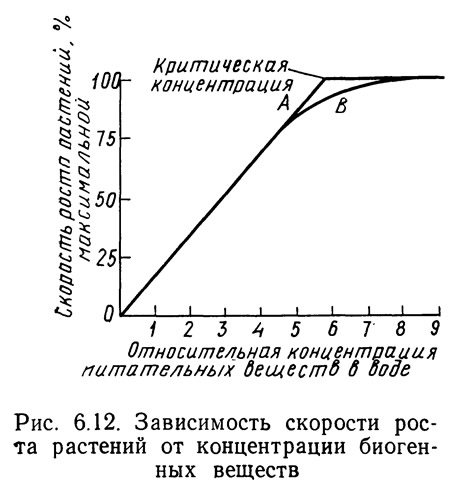
Концентрацию биогенных веществ, при которой скорость роста перестает линейно зависеть от концентрации, иногда называют критической. При концентрации выше критической может наблюдаться избыточное потребление биогенных элементов растениями, обладающими такой адаптацией. Ниже критической концентрации избыточное потребление невозможно. Критическая концентрация фосфора не постоянна для водорослей Chlorella и Scenedesmus, а зависит от величины падающей световой энергии и температуры (Azad and Borchardt, 1970). Увеличение освещенности или температуры снижает критическую концентрацию, по крайней мере, для ограниченного диапазона освещенностей и температур. Причина такого явления до конца еще не выяснена.
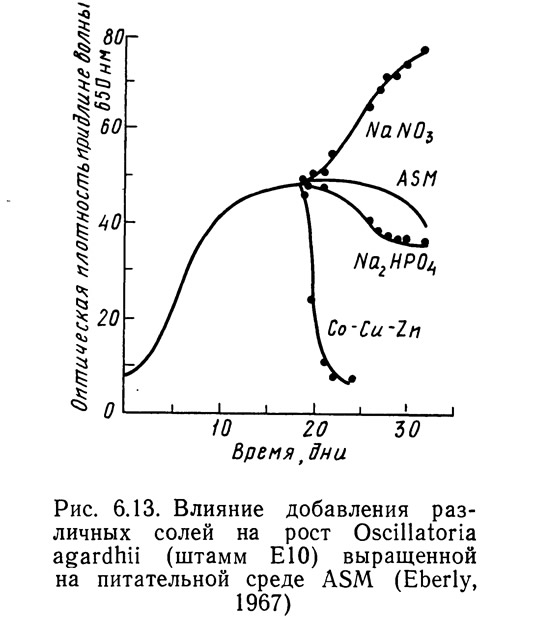
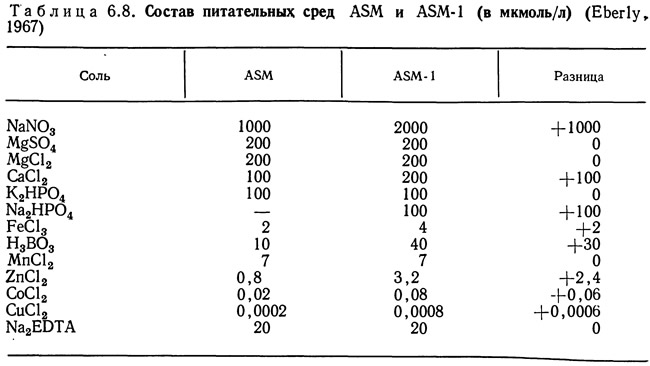
Эберли (Eberly, 1967) изучал влияние различных питательных веществ на синезеленые водоросли с использованием двух питательных сред: ASM и ASM-1 (табл. 6.8). На рис. 6.13 показано влияние изменения концентрации биогенных элементов в питательной среде на скорость роста штамма ЕЮ синезеленых водорослей Oscillatoria agardhii. Этот штамм был выделен из вод оз. Маклиш в штате Индиана. Основной питательной средой была ASM, в которую по одной добавляли различные соли в количествах, необходимых для достижения концентрации этого вещества в среде ASM-1. По вертикальной оси откладывали оптическую плотность, которая предполагалась прямо пропорциональной скорости роста водорослей. Изучаемое вещество в каждом случае добавляли на 18-й день роста растения. Увеличение концентрации кобальта, меди, цинка и Na2HPO4 замедляло рост. Добавление нитрата натрия резко увеличивало скорость роста.
Некоторым видам водорослей для поддержания роста требуются определенные витамины. К ним относится Monochrysis lutheri, которая часто используется в пищу устрицами. Между ростом клеток Monochrysis lutheri и концентрацией витамина В12 в среде существует линейная зависимость (Droop, 1961):
где n — число клеток в 1 мм; С — концентрация витамина В12, г·10-12/мл.
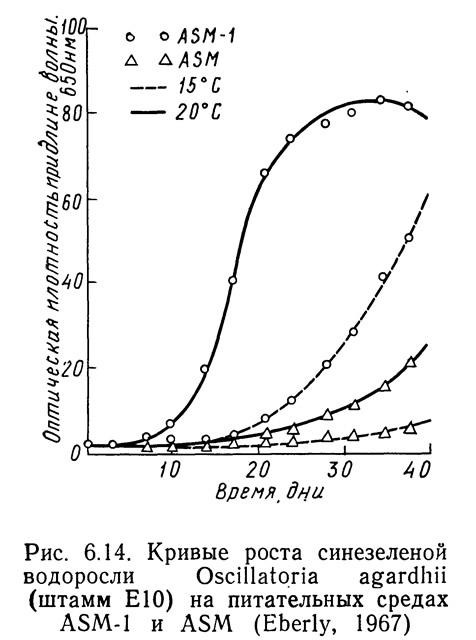
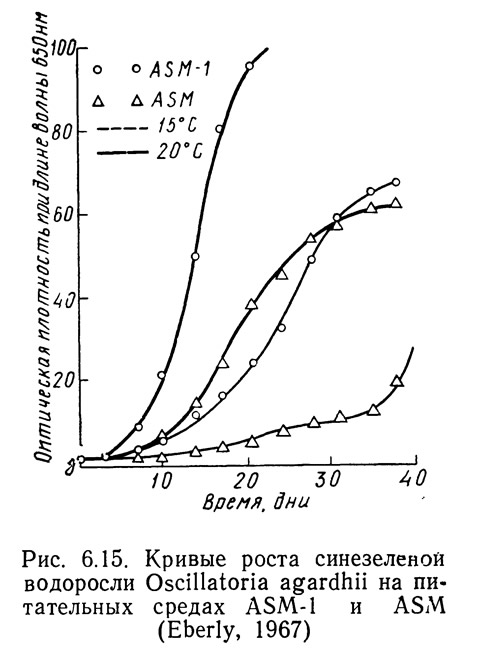
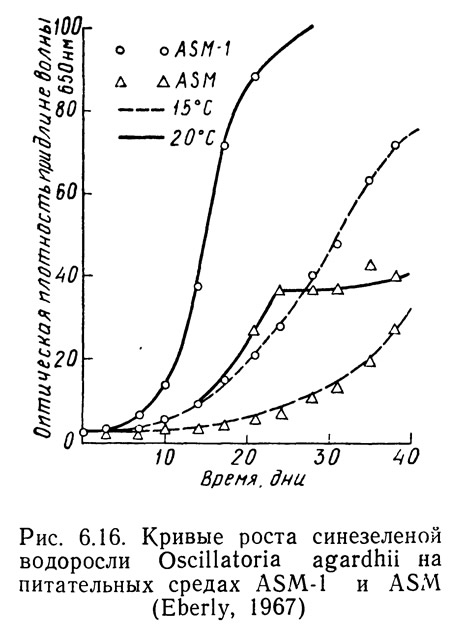
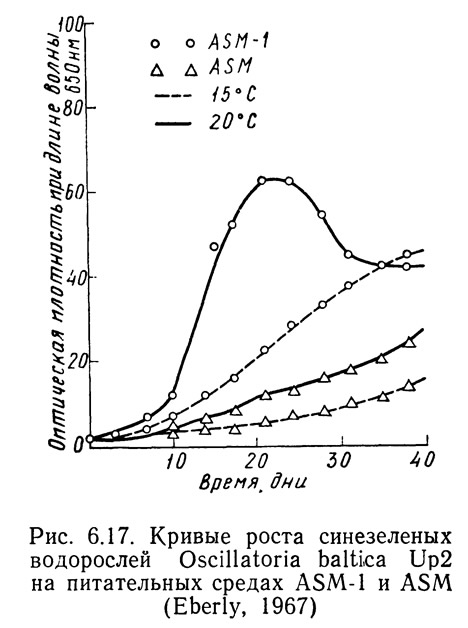
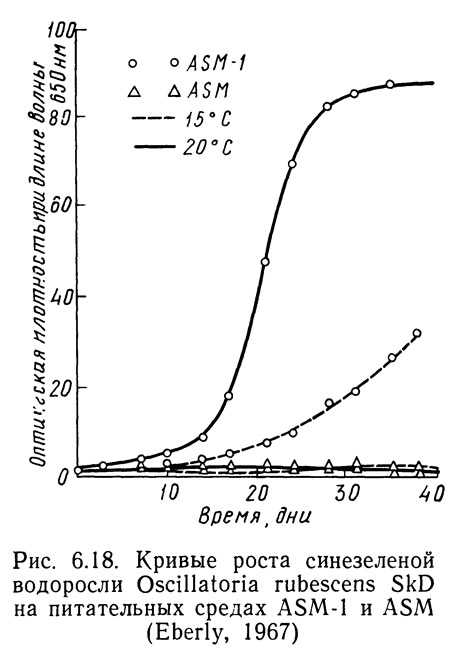
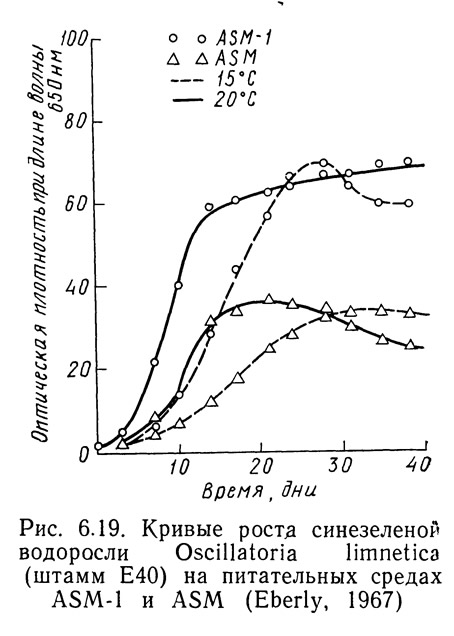
Различные штаммы одних и тех же видов могут по-разному реагировать на одинаковые условия роста в зависимости от способности к адаптации. На рис. 6.14—6.16 приведены кривые скорости роста1 Oscillatoria agardhii, взятой из разных водоемов и выращенной на среде ASM или ASM-1 при температурах от 15 до 20°С. На рис. 6.14 приведены данные для водоросли из оз. Маклиш (штат Индиана). Рис. 6.15 и 6.16 получены для водорослей из озер Гьерсьё и Акерсватн в Норвегии. На рис. 6.17 показана скорость роста Oscillatoria baltica, взятой из Балтийского моря вблизи Стокгольма (Швеция) и выращенной в средах ASM и ASM-1 при температурах 15 и 20°С. На рис. 6.18 приведены аналогичные кривые для Oscillatoria rubescens из озера Стэйнсфьорд в Норвегии, а на рисунке 6.19 — для Oscillatoria limnetica из оз. Маклиш в штате Индиана. Анализируя рис. 6.14—6.16, можно оценить влияние адаптации различных штаммов одного вида на скорости роста. Рис. 6.17—6.19 позволяют сравнить различные виды Oscillatoria.
Колебания концентрации питательных веществ (солей) оказывают дополнительное влияние на жизнь водных организмов посредством изменения осмотического давления. Высокие концентрации солей приводят к увеличению осмотического давления. Поддержание солевого баланса на поверхности организма при наличии разности осмотического давления в воде и тканях требует дополнительных затрат энергии. Если энергетические затраты слишком высоки, организм не в состоянии поддерживать необходимый солевой баланс и погибает. Таким образом, осмотическое давление или содержание солей определяет диапазон солености, в котором организм может существовать.
Примечания
1. Скорость роста измеряли по оптической плотности при длине волны 6500 А.