
Зооксантеллы сравнительно легко можно получить в культуре в виде свободно живущих водорослей. Для получения их в культуре зооксантеллы отмывают от тканей коралла раствором антибиотика и культивируют на синтетической среде или на морской воде с добавкой биогентов (Zahl, Mcl.aughlin, 1957; МсLaughlin, Zahl, 1966). В качестве источника азота в эту среду добавляют мочевину и в качестве источника фосфора-глицерофосфат и нуклеотиды. Зооксантеллы усваивают также органические кислоты (Taylor, 1973). На обедвенной питательной среде в культурах зооксантелл появляются подвижные формы, сходные с клетками планктонных динофлагеллят из рода Gymnodinium. В последнее время показано, что появление подвижных клеток связано с освещенностью. При нормальном фотопериоде в середине дня часть неподвижных клеток зооксантелл приобретает подвижность (Lerch, Cook, 1984). Исследования зооксантелл в культурах выявили у них достаточно сложный жизненный цикл. Зооксантеллы, живущие в клетках кораллов, являются одной из стадий их жизненного цикла (рис. 12.1), а именно одной из двух разновидностей ее, способных к делению неподвижных цист (McLaughlin, Zahl, 1966; Steele, 1975; Schoenberg, Trench, 1980). Они расположены поодиночке или группами по 2-3 экз. внутри клеток полипа (Kawaguti, 1964). В клетках хозяина они размножаются делением пополам. В культурах зооксантеллы находятся в виде следующих клеточных форм:
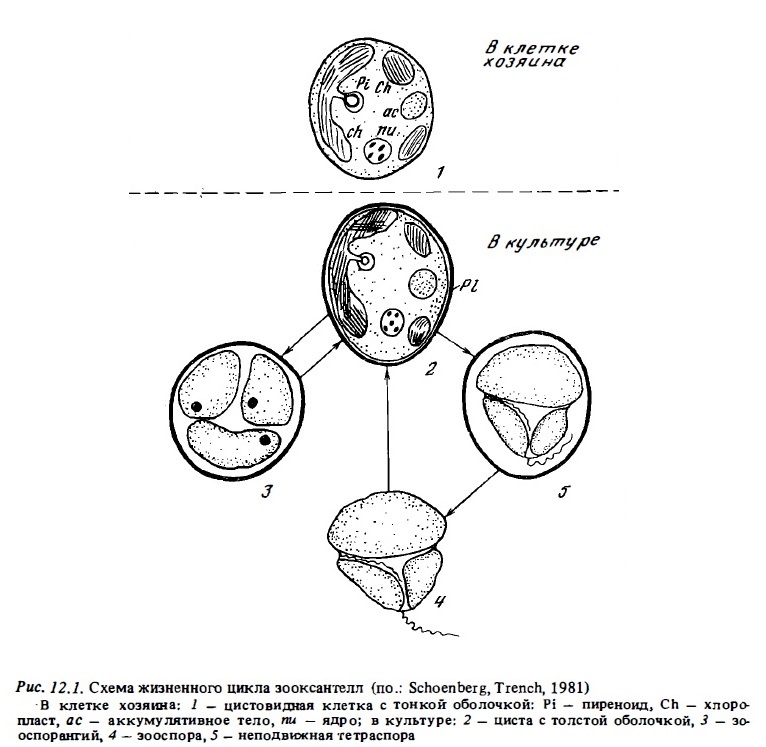
а) в виде неподвижных цист с более прочной оболочкой, чем у симбиотических их клеток-цист, живущих в тканях полипа-хозяина,
б) в виде подвижных жгутиковых динофлагеллят, сходных с планктонными гамноданиумами,
в) в виде зооспор. Неподвижные их клетки-цисты размножаются в культурах делением пополам или делением на 4 клетки - неподвижные тетраспоры.
Кроме того, часть клеток превращаются в зооспорангии, которые содержат одну крупную подвижную зооспору с двумя жгутиками, сходную по строению с планктонными гимнодишумами (см. рис. 12.1). Она активно плавает и способна внедряться в моло-днге полипы кораллов и в ткани других симбионтов рифа (Schoenberg, Trench, 1980; Trench, 19881).
У всех симбионтных полипоидных кишечнополостных зооксантеллы находятся внутри клеток зктодермы тела полипа, щупалец, орального диска и мезентерия (Vandermeulenet al, 1972). У тридакн зооксантеллы находятся в полостях кровеносных сосудов мантии.
Как мы видели выше, зооксантеллы живут в отношениях эндосимбиоза в клетках и тканях сотен разных видов, сильно различающихся по своим иммуно-генетическим и физиологическим свойствам беспозвоночных, таких, как медузы, актинии, гексакораллы, октокораллы, моллюски-тридакны (Taylor, 1969; Muscatine, 1974). При этом отдельные их штаммы сходны по своим основным систематическим признакам: размерам клеток, строению, набору пигментов, по стадиям жизненного цикла. Это дает основание относить их к одному и тому же виду динофлагеллят Symbiodinium microadriaticum. Столь широкий набор хозяев у одного вида эндосимбионта сам по себе является беспрецедентным биологическим феноменом (Schoenberg, 1980; Schoenberg, Trench, 1980). Более того, последними авторами было доказано, что один какой-либо хозяин, например, актиния Aiptasia, после выброса своих зооксантелл может вновь инфицироваться их штаммами, взятыми не только из особей того же вида, но выделенных из совершенно отличных по систематическому положению животных, например из кораллов или из тридакн, хотя сама по себе скорость инфицирования этих актиний "своим" штаммом зооксантелл идет значительно быстрее, чем "чужими" штаммами. Тем не менее появились данные, которые показывают, что зооксантеллы, полученные из разных животных-хозяев и малоразличимые морфологически, все же образуют генетически осособленные штаммы, которые достаточно четко различимы по ряду биохимических показателей, таких, как скорость электрофореза белков, состав жирных кислот и стеролов, а также по некоторым физиологическим показателям, таким, как подвижность и скорости роста в культурах in vitro, способ адаптации к недостатку света, способность инфицировать молодые особи хозяина (Kinzie et al, 1977; Schoenberg, 1980; Chang et al., 1983; Blank, Trench, 1985; Colley, Trench, 1985). Эти различия говорят в пользу того, что штаммы симбиотических зооксантелл из разных хозяев должны различаться и по своему генотипу. Последний вывод был подтвержден недавними исследованиями структуры хромосомного аппарата ядер зооксантелл, выделенных из разных по систематическому положению животных: из медузы Cassiopeia, из актиний Heteractis и Anthopleura и из коралла Montipora verrucosa. Оказалось, что морфологически малоразличимые зооксантеллы этих животных-хозяев различаются по числу хромосом и по их объему настолько, что могут быть отнесены к разным видам (Blank, Trench, 1985). В частности, зооксантеллы медузы содержали 97 хромосом, зооксантеллы актиний 74 и 50 хромосом и зооксантеллы коралла — 26 хромосом.