Таксономическое разнообразие популяций рифовых рыб в разных географических зонах оценивают, сравнивая число видов, населяющих однородные их биотопы, скажем патч-рифы (a-разнообразие). Его показателем может до известной степени служить общее число видов в популяции (S). Однако более полно оно отражается логарифмом отношения числа особей к величине S (№, Gladfelter et al., 1980) - или индексом Шеннона-Винера (Smith, 1978). Результаты сравнения ихтиофауны сходных биотопов разный географических зон выявили значительное их разнообразие даже при больших колебаниях общего числа видов (S). Так, величина № мало различалась на патч-рифах и на лагунных рифах Карибского бассейна и Тихого океана, несмотря на то что общее число заселяющих их видов рыб различается в 1,5—2 раза (Gladfelter et аl., 1980). Она выражалась величинами 0,68 на атолле Эниветок и 0,73 на карибских рифах Сан-Круа. Индекс Шеннона-Винера, равно как и таксономическое разнообразие, выражается близкими величинами на патч-рифах Багамских островов в Атлантике и островов Общества в Тихом океане: 3,4-5,2 и 2,4-5,0 соответственно (Smith, 1978). Эти данные достаточно однозначно свидетельствуют о закономерном характере формирования популяций рифовых рыб, о значительной степени их предсказуемости. Их состав определяется характером видоспецифических адаптаций наличной ихтиофауны рифа, биологическими ресурсами рифа и пространственной структурой биотопов. Опыта ряда авторов показали, что при заселении отдельный патч-рифов, ихтиофауна которых была предварительно уничтожена ротеноном, изменения ее состава оказывались минимальными — она практически полностью восстанавливалась в прежнем виде (Smith, 1977), что указывает на детерминированность структуры сообщества рифовых рыб (теория "порядка": Smith, Tyler, 1972; Gladfelter et al., 1980).
Последняя точка зрения оспаривается некоторыми авторами (Sale, 1978, 1980; Talbot et аl., 1978), которые считают, что структура сообществ рифовых рыб носит стохастический характер (теория "хаоса") и может значительно варьировать в сходных биотопах, скажем на разных патч-рифах, что делает ее малопредсказуемой. В соответствии с представлениями сторонников теории "хаоса" в биотопах рифа ввиду высокой смертности от выедания и других причин постоянно освобождается часть пространства для заселения его оседающей молодью, которая рекрутируется из состава наличных в данное время и случайно подобранных пелагических личинок. Детальный анализ обоих точек зрения с позиции теории оборота состава стай рыб (Bohnsack, 1983) показывает, что вывод о малой предсказуемости, будучи следствием слишком малого временного шага, является в значительной степени артефактом. Более длительные наблюдения за сообществами одного и того же рифа, а также сравнения их состава в сходных по строению и зональности биотопах подтверждают детерминистический характер структуры сообществ рифовых рыб, т.е. теорию "порядка" (Thresher, 1977). Эту теорию подтверждают результаты длительных наблюдений за сообществом рыб, заселивших большой искусственный риф, который был сооружен Рэнделлом в 1960 г. на о-ве Сент-Джон (Виргинские острова). Уже через два месяца на нем сформировалось сообщество рыб, которое в последующие годы подвергалось относительно небольшим изменениям по таксономическому составу и по общей численности особей. Так, в 1962 г. на нем обитало 39 видов из 14 семейств. В 1979 г. на этом же рифе обитало 40 видов из 14 семейств при сохранении общего уровня разнообразия и численности доминирующих семейств. Последние располагались го этому признаку в следующем порядке: Pomadasysiidae, Holocentridae, Serranidae, Acanthuridae, Labridae, Scaridae, Po macentridae, Chaetodontidae, Apogonidae, Muraenidae, Clinidae, Cirrithidae, Tetraodonti-dae (Ogden, Ebersole, 1981).
Детерминированный характер структуры сообществ рифов рыб обусловлен высокой степенью их специализации в отношении использования разнообразный ресурсов рифа и наличием у них тонких видоспецифических адаптаций. Этот фактор оказывает определяющее влияние на дальнейшее выживание рекрутированной молоди и обеспечивает однотипность и относительное постоянство сообществ рыб в сходных по типу- биотопах в течение длительного времени (Smith, 1978; Anderson et аl., 1981). Важнейшими факторами, влияющими на состав сообществ рыб и их численность, являются в этих условиях, во-первых, пространство и морфологическая структура каждого биотопа, которые определяют наличие возможных убежищ (Thresher, 1977; Sale, 1978; Greenfild, Greenfield, 1982), и, во-вторых, кормовые ресурсы биотопа (Wiliiams, Hatcher, 1983). В целом можно признать, что сообщества рыб коралловых рифов формируются в соответствии с теми же принципами относительной детерминированности их таксономической и возрастной структуры, как и сообщества наземных позвоночных (Anderson et al., 1981). Такая детерминированность не исключает элементов стохастичности в формировании сообществ отдельных участков рифа и в их сезонных изменениях. Одна из причин таких изменений состоит в том, что наряду со специализированными видами рыб, подбор которых зависит в значительной степени от характера биотопа, они включают также менее специализированные виды, которые менее требовательны в отношении условий среда. Они обладают широким пищевым спектром и могут перекрывать друг друга в использовании пространства и ресурсов рифа. Их рекрутирование и вносит определенный элемент стохастичности в структуре популяции рифовых рыб, отмеченный автором теории "хаоса" (Sale, 1978). Интенсивное выедание также вносит элемент стохастичности в формирование сообществ рифовых рыб, обусловливая неравномерную и достаточно высокую оборачиваемость видов в отдельных биотопах рифа (Talbot et аl., 1978).
Сообщества рыб разнотипные биотопов, расположенных в разных зонах одного и того же рифа, различаются по своему составу значительно больше, чем однотипные биотопы разных рифов (Hiatt, Strasbourg, 1960; Hobson, 1974; Smith, 1978; Williams, Hatcher, 1983). Состав сообществ рыб в отдельных биотопах рифа во многом определяется их строением, глубиной и зональным положением в геоморфологической структуре рифа (Gosline, 1971; Chave, Eckert, 1974; Jones, Chase, 1975; Goldman, Talbot, 1976; Smith, 1977). Большинство рифовых рыб обитает только в пределах определенных структурных элементов рифа (Sale, 1978а). Это положение можно продемонстрировать на следующих примерах. Для 8 разных биотопов рифа Уантри (Б. барьерный риф) общими были только 7% всего числа обнаруженных на них видов рыб. В то же время около 50% видов рыб из общего их числа, обнаруженного в каждом из этих биотопов, обитали только на данном биотопе и не встречались в остальных 8 биотопах (Goldman, Talbot, 1976). При анализе состава ихтиофауны в 7 разных биотопах атолла Фаннинг бьто обнаружено, что в среднем только 26% видов рыб были общими для двух соседних биотопов от их суммарного числа в обеих биотопах (Chave, Eckert, 1974). Для рифа Бимини этот процент был близким к 34% (Clarke, 1977).
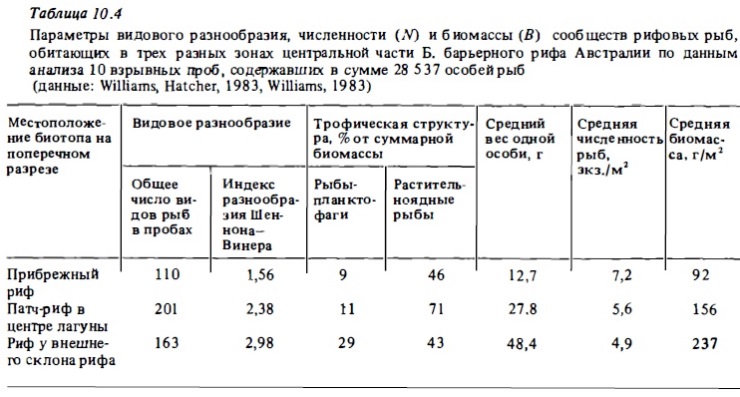
Исследования, выполненные на Б. барьерном рифе (Williams. 1983), показали, что состав сообществ рыб, населяющих разные зоны рифа на поперечных разрезах от берега до внешнего склона, достаточно сильно варьирует (табл. 10.4). Эти изменения состава ихтиофауны в разных биотопах одного и того же рифа оказались намного более значимыми, чем его вариации в сходных биотопах разных участков Б. барьерного рифа, расположенных между 11 и 22° ю.ш. Около 60% видов из их общего числа (323 вида) были встречены только на одном из трех рифов, располагавшихся в разных зонах поперечного разреза, и только 7% видов были встречены на всех трех рифах (Williams, 1983). Видовое разнообразие сообществ рыб возрастало по направлению от шельфа к внешнему склону. Часто значительные изменения видового состава и численности рыб можно было наблюдать по периметру кольцевых рифов. Эти изменения были связаны с различиями в составе кораллов, заселяющих разные участки кольцевого рифа, а также с разной интенсивностью прибоя (Done, 1982; Williams, 1982).
Приведенные выше данные показывают, что сообщества рифовых рыб, населяющих разные биотопы, характеризуются высокой степенью дискретности и практически не смешиваются между собой, будучи привязаны к определенным структурным элементам и зонам рифа. Такая оседлость характерна только для популяций рифовых рыб и, как правило, не свойственна популяциям прибрежной зоны внерифовой ихтиофауны. Большинство рифовых рыб и их видовых ассоциаций не меняют своих постоянных мест обитания на рифе. В этих условиях при очень высокой плотности популяций каждый элемент субстрата рифа имеет своего хозяина (Smith, 1978). Поэтому можно полагать, что главным фактором, лимитирующим общее число видов рыб и их численность в отдельных зонах рифа, является в первую очередь пространство, которое в данном случае важнее, чем кормовые ресурсы или выедание. От размеров пространства и от структуры донных биотопов рифа зависят наличие убежищ и гнезд, условия охоты и размножения.
Выше было показано, что рыбы, обитающие на рифе, не образуют единого сообщества, а разделяются на несколько ассоциаций, экологически обособленных в пределах отдельных биотопов и зон рифа, где они занимают доминирующее положение. Эти ассоциации так или иначе связаны между собой и взаимодействуют друг с другом в отношении использования ресурсов рифа (Vivien, 1973). При этом часть рыб с ярко выраженным территориальным поведением постоянно живет в зоне флета или на внутренних патч-рифах. В их числе многие мелкие донные рыбы, такие, как Pomacentridae, Gobiidae. Другая часть популяции таких рыб - это молодь более крупных хищников рифа, таких, как груперы или губаны, взрослые особи которых живут преимущественно в зоне внешнего склона. Эти молодые рыбы прячутся в убежищах флета и патч-рифов от выедания. Они временно входят в состав популяций зоны флета и патч-рифов и ими эксплуатируются. Отмечено, что видовое разнообразие рыб отдельных биотопов рифа зависит от сложности структуры их твердого субстрата (Hobson, 1974; Hatcher, 1981). Например,в зонах рифа, занятых живыми ветвистыми кораллами, общее число видов рыб обычно бывает значительно выше, чем в зонах рифа, обедненньх кораллами (Harmelin — Vivien, 1977; Carpenter et аl., 1981).
Ввиду относительного постоянства ресурсов пространства и пищи локальные ассоциации рифовых рыб и даже отдельные их стаи имеют относительно стабильный состав. Таковы, например, стаи рыб-попугаев или рыб-хирургов. При этом резиденты, которые занимают постоянное место обитания, имеют преимущество перед вселенцами активно их атакуют и прогоняют. Если резидент погибает, его место занимает другая рыба, но не всегда того же вида (Sale, 1974). Самые строгие резиденты рифа, живущие только в определенном виде субстрата, обеспечивают стабильность состава ихтиофауны сходных биотопов. В их числе представители семейств: Pomacent ridae, Eleotridae, Gobiidae, Muraenidae, Scorpaenidae. Это так называемые стержневые вида сообщества ("core species"). Менее строгие резиденты рифа, такие, как губаны, груперы, рыбы-флейты, рыбы-попугаи, рыбы-хирурги, могут использовать в качестве убежищ разные виды субстрата. Они создают разнообразие состава ихтиофауны однотипных биотопов разных рифов. Существенно, что в регионах с меньшей общей численностью видов на рифе количество таких "общих" для разных биотопов видов возрастает. В результате в регионах с меньшей общей численностью видов рыб на всем рифе общая численность особей в популяциях на сходных по типу биотопах может быть в них соизмеримой, хотя, казалось бы, при более высоком разнообразии следовало ожидать и более высокой численности за счет более полного использования ресурсов рифа. Такие данные были получены, например, при анализе тотальных проб рыб, собранных после глушения, на тихоокеанском рифе Уантри и на восточноафриканском рифе Тутиа, где общее число видов рыб на рифе было в 2,5 раза меньше, чем на первом (Goldman, Talbot, 1976). Эти данные говорят в пользу того, что в ходе эволюции ихтиофауны рифов появилось "избыточное" количество однотипных по своей трофической характеристике видов рыб, которые могут взаимно заменять друг друга в разных биотопах. Не случайно поэтому в пределах одного рифа общее число видов варьирует в разных его зонах в большей степени, чем абсолютная численность популяций. Так, на барьерном рифе Кокос (о-в Гуам) общая численность рыб варьировала в пределах 11-17 экз./10 м2 при измерении общего числа видов в 5 раз (Jones, Chase, 1975).
Наибольшее видовое разнообразие по показателю общего числа видов отмечено в сообществах, обитающих у внешних склонов рифа и на флете, т.е. в зонах с наиболее интенсивным водообменом. Высокое видовое разнообразие рыб в этих зонах связано, по-видимому, главным образом с тем, что здесь обитает много видов мелких рыб из сем. Pomacentridae и Gobiidae, питающихся зоопланктоном, который приносится с проходящими над рифом водами океана (Hobson, Chess, 1978; Hannelin-Vivien, Bouchon-Navaro, 1981). Здесь же концентрируются хищные виды, которые охотятся за этими мелкими планктофагами, а также растительноядные рыбы, которые в период высокой воды уходят с внешней стороны рифа кормиться на флет.