
По своему внешнему виду зооксантеллы представляют собой круглые или слегка овальные буро-зеленые водорослевые клетки. Размеры зооксантелл варьируют от 8 до 14 мкм. При этом их штаммы, которые заселяют хозяев далеко отстоящих друг от друга таксонов могут значимо отличаться по своим размерам (Blank, Trench, 1985). Средние размеры зооксантелл большинства кораллов близки к 9-11 мкм. Согласно Смиту, зооксантеллы находятся преимущественно внутри клеток эндодермы и гастродермы полипа в полостях их вакуолей (Smith, 1979). Часть зооксантелл локализована также в межклеточниках (Воскобойников и др., 1983). Они отделены от клетки хозяина плотной оболочкой-перипластом, которая имеет сложное строение и состоит минимум из трех мембран, имеющих в своей структуре целлюлозу (McLaughlin, Zahl, 1966; Taylor, 1973; Muscatine, 1974; Mashansky et al., 1980). Функциональное значение этой внешней оболочки зооксантелл очень велико. Она надежно изолирует ее от атаки лизосом хозяина, предотвращая ее переваривание (Muscatine et al., 1975). В то же время она проницаема дня обмена метаболитами с клеткой хозяина. На некоторых ее участках имеются многочисленные выпячивания и отслоения наружной мембраны. Предполагается, что таким путем происходит частичная транслокация продуктов фотосинтеза зооксантелл (в виде капелек жира) в клетки хозяина (Patton, Burris, 1983). Выросты клетки полипа образуют своего рода капсулу вокруг зооксантеллы (Воскобойников и др., 1983). Неподвижные клетки-цисты в культурах зооксантелл отличаются от симбиотических их клеток, находящихся в тканях хозяина тем, что имеют еще более прочную и толстую оболочку (Schoenberg, Trench, 1980). Она предохраняет их 6т переваривания при "заражении" новых хозяев через содержимое кишечников животных — их жертв.
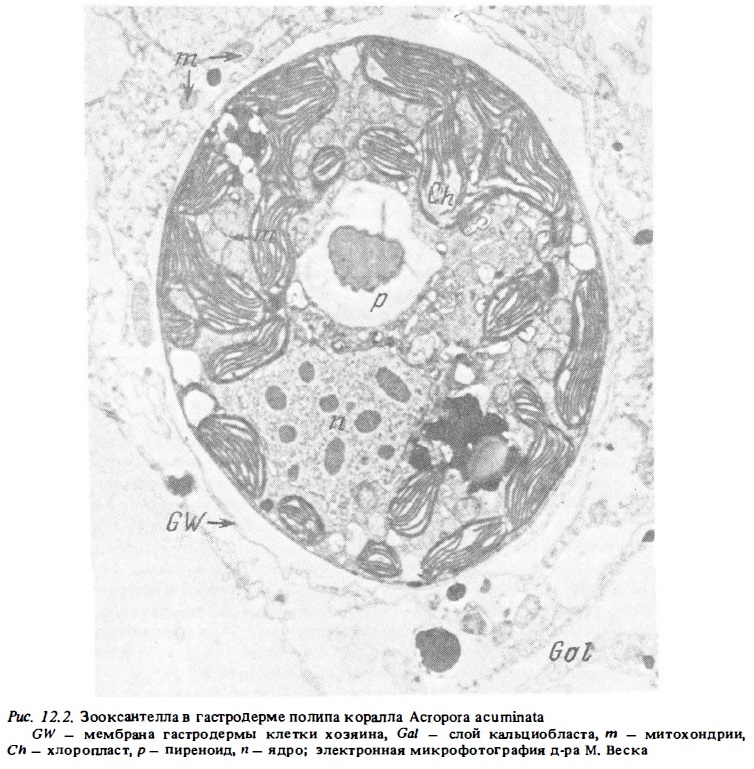
Наличие перипласта снижает различие между внеклеточными межорганизменным симбиозом и эндосимбиозом (Smith, 1979). Действительно, находясь внутри клеток хозяина, зооксантеллы в значительной мере изолированы от нее перипластом. Они сохраняют способность независимого развития, о чем говорят успешное их культивирование и прохождение ими стадий развития вне клеток хозяина. После внедрения зооксантеллы в клетку хозяина между ней и клеткой хозяина устанавливается обмен метаболитами. Величина его потоков контролируется последней наряду с контролем темпов размножения зооксантелл в клетках хозяина и их численности. Численность зооксантелл в клетках хозяина регулируется путем выброса избыточных их клеток из тканей полипа наружу (Steele, 1976). Другой механизм регулирования их численности состоит в ограничении поставки хозяином в зооксантеллы необходимых им биогенов (Reimer, 1971; Drew, 1972). Примечательной чертой строения клеток зооксантелл является наличие в них крупного хлоропласта, занимающего почта половину всего их объема (рис. 12.2). Хлоропласт зооксантелл расположен по периферии клетки. Он имеет дольчатое строение. Его ламеллярная система состоит из 10-12 комплексов, в каждом из которых упаковано по 3 тилакоида. Периноид хлоропласта крепится к нему с помощью ножки. В клетке зооксантеллы имеется ядро и так называемое аккумулятивное тело. Последнее имеет вид крупной вакуоли, наполненной рыхлым материалом (Singh, Merser, 1976; Schoenberg, Trench, 1980; Воскобойников и др., 1983).
Первоначальное "заражение" полипов зооксантеллами происходит по-разному у разных видов кораллов и других Anthozoa и Hydrozoa. Зооксантеллы присутствуют в яйцах или личинках (планулах) гидроидов и гидрокораллов (Agalophenia, Millepora), актиний (Anthopleura), октокораллов (Xenia, Eunicella), а также в планулах многих склерактаний, таких, как Agaricia. agaricites, Siderastrea radians, Pocillopora damicornis, Stylophora pistillata, Galaxea aspera. Зооксантеллы встречаются и в яйцах нерестующих кораллов; Fungia actiniformis, Favia fragum (Marshall, 1932; Gohar, 1940; Atoda, 1951; Smith, 1980; Fitt, 1984). Находясь в клетках планул, зооксантеллы активно фотосинтезируют, снабжая их энергией. Таким образом ОНИ способствуют лучшему их выживанию в планктонной их стадии (Richmond, 1981). В то же время многие виды морских симбионтных животных продуцируют яйца или рождают личинки, в которых изначально зооксантеллы отсутствуют (Atoda, 1953; Droop, 1963; Kinzie, 1974; Trench, 1980; Fitt, 1984, Babcock et. al., 1986). В их числе сцифомедузы (Cassiopeia и Mastigias), октокораллы (Heteroxinia, Briareum, Pseudopterogorgia), преобладающая часть нерестующихся склерактиниевых кораллов, а также симбионтные моллюски-тридакны. Личинки симбионтных беспозвоночных, лишенные изначально зооксантелл, "зараждаются" ими, будучи уже в морской среде. В опытах Кинзи (1974) новорожденные планупы горгонарии Pseudopterogorgia без наличия зооксантелл осаждали на субстрат в аквариумах с водой, лишенной водорослей. Когда из осевых планул сформировались молодые полипы, в аквариум добавляли взвесь клеток разных штаммов динофлагеллят, в том числе свободно живущих типа гимнодиниумов, и зооксантеллы,выращенные в культуре, часть клеток которых находилась в подвижной стадии. Наблюдения показали, что динофлагелляты, включая свободно живущие, привлекались молодыми полипами, концентрировались вблизи них и заплывали им в рот. "Заражение" же полипов происходило только зооксантеллами Symbiodinium microadriaticum.
Локация полипов свободно живущими в планктоне подвижными зооспорами зооксантелл происходит при посредстве хеморецепции. Зооспоры двигаются в сторону повышенной концентрации солевого азота, выделяемого полипами кораллов и другими беспозвоночными. Они попадают в кишечную полость или в кровоток беспозвоночных не только в свободном состоянии, но и в составе содержимого кишечников планктонных рачков, которых заглатывают животные — их потенциальные хозяева (Fitt, 1984). Будучи окружены плотной оболочкой, зооксантеллы долго не перевариваются в кишечнике рачка. Таким путем после его заглатывания зооксантеллы попадают в кишечную полость и инфицируют клетки своего потенциального хозяина (Schoenberg, 1980). Сам процесс инфицирования тканевых клеток зооксантеллами, попавшими внутрь мезентерия полипа,. происходит путем их фагоцитоза на мезентериальных нитях (Kinzie et al., 1977; Fitt, Trench, 1980). Источником зооксантелл в водах рифа при заражении ими планул в естественных условиях может быть их выброс в воду кораллами и тридакнами (Goreau, 1964), а также их поступление при выедании кораллов ежами и особенно рыбами-попугаями, которые обнажают и разрушают ткани полипов, освобождая зооксантеллы (Fitt, Trench, 1980). Зооксантеллы, потребленные кораллофагами вместе с полипами и слизью, плохо ими перевариваются. В их фекалиях в большом количестве присутствуют живые зооксантеллы, способные заражать своих потенциальных хозяев, тем более что полипы кораллов актиний и гидроидов охотно заглатывают фекалии рыб (Parker, 1984). Имеются сообщения об обнаружении значительного количества дино-флагеллят, неотличимых от подвижных форм зооксантелл в водах мелководий рифа (Taylor, 1980).
Находясь уже в клетке хозяина, зооксантелла может избегнуть переваривания, если она будет "узнана" клеткой хозяина как его симбионт. В противном случае зооксантелла должна иметь защиту от возможного переваривания. Это можно было бы выяснить, определив способность хозяина "узнавать" своих эндосимбионтов. Однако сведения на этот счет пока скудны. Второй механизм-защита клеток зооксантелл — более очевиден, поскольку, как отмечалось выше, они имеют защитную оболочку, которая препятствует атаке лизосом клетки хозяина (Muscatine et аl., 1975). Избыточные зооксантеллы выбрасываются клеткой хозяина, не подвергаясь перевариванию, или подвергаются автолизу (Schenberg, Trench, 1980). Выброс здоровых и отмирающих избыточных клеток идет одновременно с секрецией слизи, которая, как правило, содержит много клеток зооксантелл. Выброс зооксантелл полипами усиливается при неблагоприятных условиях температуры, освещения солености (Yonge, Nicholls, 1931; Goreau, 1964; Taylor, 1973).
- Главная
- Природа
- Коралловые рифы
- Строение зооксантелл, способы инфицирования хозяев
Подразделы категории
Поделитесь с друзьями
Похожие статьи
Фотосинтетическая функция зооксантелл
Природа » Коралловые рифы
Метаболические взаимодействия зооксантелл с полипом-хозяином
Природа » Коралловые рифы
Баланс питания кораллов и удельная скорость роста полипов и зооксантелл
Природа » Коралловые рифы
Строение и работа пищеварительной системы карпа
Водоемы » Корма для рыб
Строение и работа пищеварительной системы форели
Водоемы » Корма для рыб
Способы дезинфекции воды
Водоемы » Фильтрация воды
Питание склерактиниевых кораллов: способы захвата пищи и ее переваривание
Природа » Коралловые рифы
Способы изготовления рыбной муки
Водоемы » Корма для рыб
Способы расчета состава рецептов комбикормов
Водоемы » Корма для рыб
Технологические способы изготовления комбикормов
Водоемы » Корма для рыб
Способы повышения переваримости кормов
Водоемы » Корма для рыб
Способы подготовки сырья для комбикормов: термическая обработка конвективным методом
Водоемы » Корма для рыб
Способы подготовки сырья для комбикормов: термическая обработка кондуктивным методом
Водоемы » Корма для рыб
Способы подготовки сырья для комбикормов: тостирование
Водоемы » Корма для рыб
Природа » Коралловые рифы
Метаболические взаимодействия зооксантелл с полипом-хозяином
Природа » Коралловые рифы
Баланс питания кораллов и удельная скорость роста полипов и зооксантелл
Природа » Коралловые рифы
Строение и работа пищеварительной системы карпа
Водоемы » Корма для рыб
Строение и работа пищеварительной системы форели
Водоемы » Корма для рыб
Способы дезинфекции воды
Водоемы » Фильтрация воды
Питание склерактиниевых кораллов: способы захвата пищи и ее переваривание
Природа » Коралловые рифы
Способы изготовления рыбной муки
Водоемы » Корма для рыб
Способы расчета состава рецептов комбикормов
Водоемы » Корма для рыб
Технологические способы изготовления комбикормов
Водоемы » Корма для рыб
Способы повышения переваримости кормов
Водоемы » Корма для рыб
Способы подготовки сырья для комбикормов: термическая обработка конвективным методом
Водоемы » Корма для рыб
Способы подготовки сырья для комбикормов: термическая обработка кондуктивным методом
Водоемы » Корма для рыб
Способы подготовки сырья для комбикормов: тостирование
Водоемы » Корма для рыб
Строение зооксантелл, способы инфицирования хозяев
Дата: Февраль 2017 года
ПРЕДЫДУЩИЕ СТАТЬИ
Таксономия и специфичность водорослей-симбионтов зооксантил
Природа » Коралловые рифы
Биомасса живой ткани кораллов и ее структурных элементов
Природа » Коралловые рифы
Особенности экологической морфологии кораллов
Природа » Коралловые рифы
Вегетативное размножение кораллов
Природа » Коралловые рифы
Половое размножение кораллов
Природа » Коралловые рифы
Факторы, определяющие структуру сообществ кораллов в разных зонах рифа
Природа » Коралловые рифы
Восьмилучевые кораллы как компонент сообществ рифа
Природа » Коралловые рифы
Зоантарии в сообществах коралловых рифов
Природа » Коралловые рифы
Таксономия и специфичность водорослей-симбионтов зооксантил
Природа » Коралловые рифы
Биомасса живой ткани кораллов и ее структурных элементов
Природа » Коралловые рифы
Особенности экологической морфологии кораллов
Природа » Коралловые рифы
Вегетативное размножение кораллов
Природа » Коралловые рифы
Половое размножение кораллов
Природа » Коралловые рифы
Факторы, определяющие структуру сообществ кораллов в разных зонах рифа
Природа » Коралловые рифы
Восьмилучевые кораллы как компонент сообществ рифа
Природа » Коралловые рифы
Зоантарии в сообществах коралловых рифов
Природа » Коралловые рифы
СЛЕДУЮЩИЕ СТАТЬИ
Фотосинтетическая функция зооксантелл
Природа » Коралловые рифы
Метаболические взаимодействия зооксантелл с полипом-хозяином
Природа » Коралловые рифы
Черты экологической физиологии фотосинтеза кораллов: суточный ход фотосинтеза
Природа » Коралловые рифы
Зависимость фотосинтеза кораллов от интенсивности светового потока
Природа » Коралловые рифы
Экологическая биохимия кораллов
Природа » Коралловые рифы
Антибиотики, токсины и простагландины
Природа » Коралловые рифы
Состав и функция слизи у кораллов
Природа » Коралловые рифы
Автотрофное питание кораллов
Природа » Коралловые рифы
Фотосинтетическая функция зооксантелл
Природа » Коралловые рифы
Метаболические взаимодействия зооксантелл с полипом-хозяином
Природа » Коралловые рифы
Черты экологической физиологии фотосинтеза кораллов: суточный ход фотосинтеза
Природа » Коралловые рифы
Зависимость фотосинтеза кораллов от интенсивности светового потока
Природа » Коралловые рифы
Экологическая биохимия кораллов
Природа » Коралловые рифы
Антибиотики, токсины и простагландины
Природа » Коралловые рифы
Состав и функция слизи у кораллов
Природа » Коралловые рифы
Автотрофное питание кораллов
Природа » Коралловые рифы
ССЫЛКА НА СТАТЬЮ В РАЗЛИЧНЫХ ФОРМАТАХ
HTMLBB CodeText
Комментарии к этой статье
Еще нет комментариев
Aquaristics.ru © 2016 • Письмо админу • Карта сайта • Поиск • Мобильная версия
Оборудование: Общая информация · Аквариумы · Вода для аквариума · Освещение · Фильтрация · Аэрация · Обогрев · Грунт и декорации
Рыбки: Общая информация · Содержание и уход · Виды рыбок · Корма для рыб · Болезни и лечение · Морские животные
Растения: Общая информация · Виды растений · Уход за растениями · Посадка растений · Болезни растений
Дизайн: Планировка · Растительность
Водоемы: Общая информация · Виды водоемов · Вода и воздух · Насосы · Фильтрация воды · Аэрация · Прочие устройства · Корма для рыб
Природа: Морские водоросли · Коралловые рифы
Рыбки: Общая информация · Содержание и уход · Виды рыбок · Корма для рыб · Болезни и лечение · Морские животные
Растения: Общая информация · Виды растений · Уход за растениями · Посадка растений · Болезни растений
Дизайн: Планировка · Растительность
Водоемы: Общая информация · Виды водоемов · Вода и воздух · Насосы · Фильтрация воды · Аэрация · Прочие устройства · Корма для рыб
Природа: Морские водоросли · Коралловые рифы