В литературе по зоопланктону коралловых рифов эти достаточно четко ограниченные типы планктеров многие авторы часто путают между собой. Следствием этого может оказаться и неверная трактовка получаемый результатов. Особенно часто смешивают понятия демерсального планктона и меропанктона. Так, Портер (Porter, 1974) и Портер и Портер (Porter, Porter, 1977) всех донных беспозвоночный, которые в ночное время поднимаются в толщу воды и улавливаются ловушкой, относят к демерсальному зоопланктону. Сюда попадает и личиночный меропланктон и прилавливаемш ловушкой голопланктон (особенно копеподы и голопланктонные мизиды. Другие авторы (Ricard et аl., 1979) называют меропланктоном всех планктеров, которые прячутся днем в субстрате дна или на его поверхности. Такое смешение понятий и особенно отнесение к меропланктону или к демерсальному планктону всего зоопланктона, улавливаемого донными ловушками (Porter et аl., 1977; Alldredge, King, 1977), может затруднить оценки трофических взаимоотношений сообществ рифа и океана.
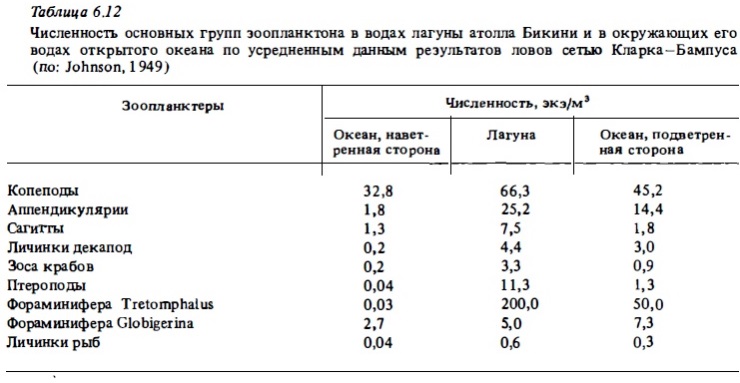
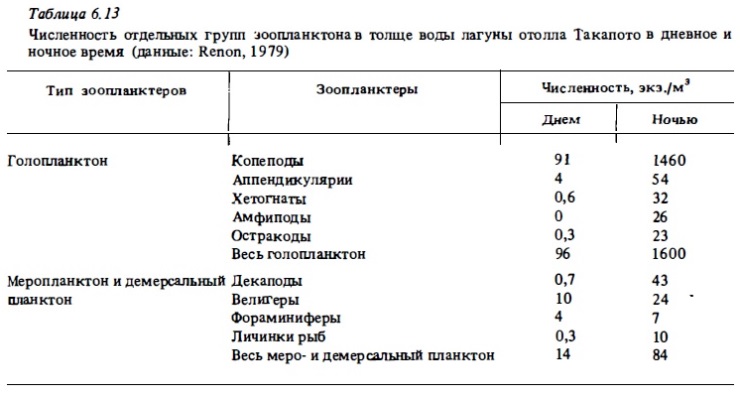
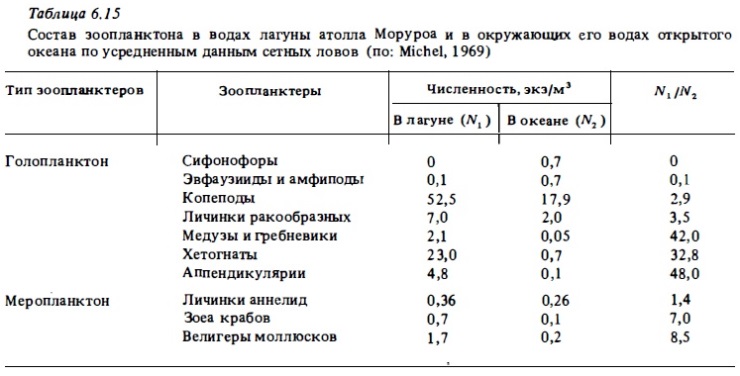
Общее число видов, регистрируемое в составе рифового планктона, варьирует у разных авторов в зависимости от применяемый орудий лова. Дневные ловы сеткой в приглубых местах рифа, особенно в зонах достаточно интенсивного водообмена с океаном, недоучитывают локальную фауну рифа, особенно демерсальные и меропланктонные ее компоненты, а учитывают в значительной степени голопланктонные виды океанского зоопланктона, приносимого на риф извне (Johnson, 1954; Gerber, Marshall, 1975; Moore, Sander, 1976; Ferraris, 1982; Goswami, 1983). При такой методике сбора число видов зоопланктона, обнаруживаемое в водах лагун, бывает близким к 100-120. При использовании ночных ловов это число значительно возрастает за счет видов, мигрирующих ночью со дна (табл.6.12, 6.13). Оно достигает максимума (до 260 видов) при использовании ловушек (Sale et al., 1976, Мс William et al., 1981). Хотя даже ловы сетью уже выявляют заметное отличие состава лагунного зоопланктона от зоопланктона океана благодаря обогащению его фауны демерсальными видами (табл.6.14, 6.15), значительные вариации состава зоопланктона получаются при использовании световых ловушек пли ловов с использованием отсасывания зоопланктеров, привлекаемых ночью светом (Emery, 1988). В таких пробах, например, вместо обычного доминирования копепод преобладают личинки крабов, амфиподу стоматоподы, полихеты, кумовые рачки, мизидь Такое же доминирование демерсальных форм зоопланктона, а также мизид и амфипод в зоопланктоне рифа было установлено нами при использовании ночных его ловов путем процеживания 150 л воды через планктонную сетку (табл. 6.15-6.18). Сейл и др. (Sale et al., 1976), пользуясь световой ловушкой для сборов зоопланктона в воде над рифами, идентифицировали в собранных пробах 106 видов (табл.6.19). При этом преобладающее их число (66 видов) встречалось только в воде над рифом и отсутствовало в пробах, полученных в прилегающих водах открытого моря. Остальные виды были общими для зоопланктона рифа и для вод открытого моря.
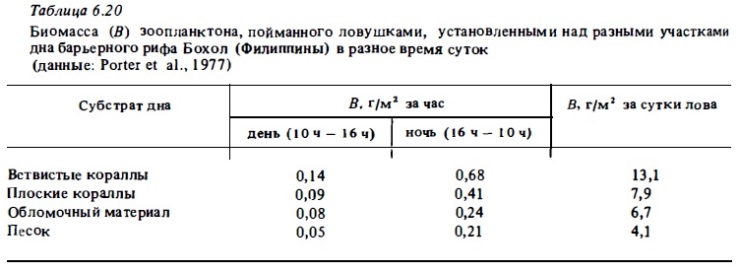
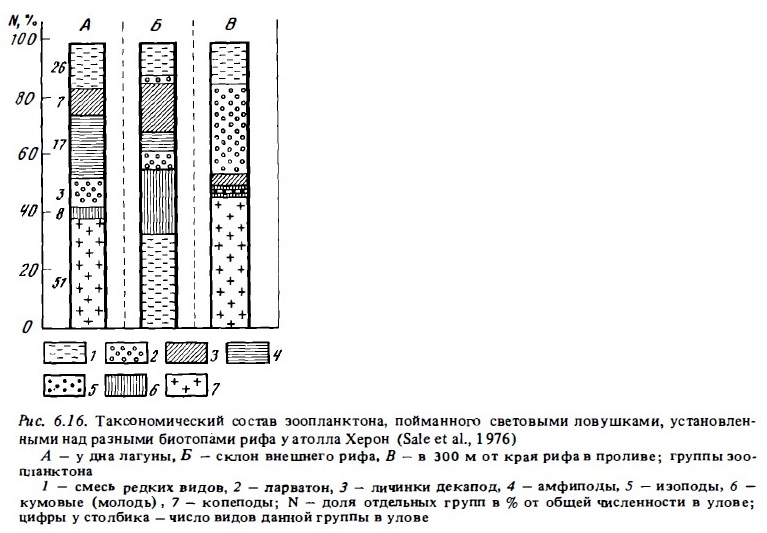
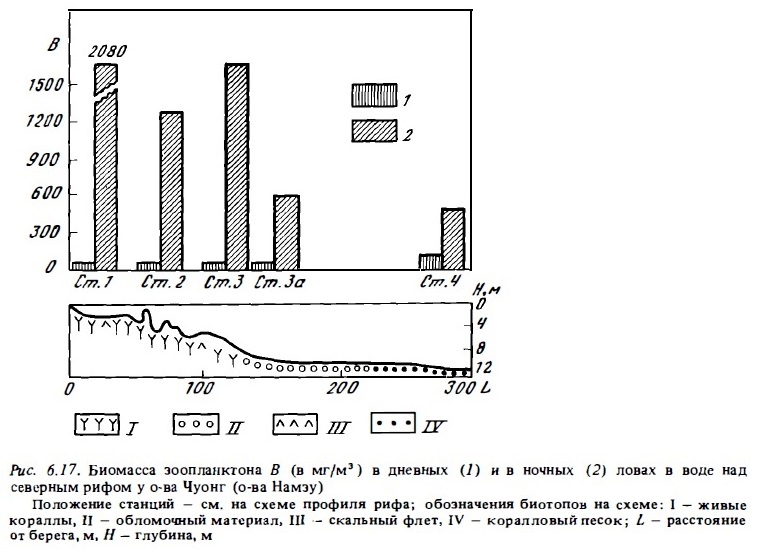
Состав фауны зоопланктона (табл. 6.20; рис. 6.16, 6.17) значительно варьирует при его сборах над разными биотопами рифа (von Hentig, 1971; Sale et al., 1976; Alldredge, King, 1977; Porter et al., 1977; Мс William et al., 1981; Birkeland, Smalley, 1981), а также в разных зонах лагуны (Sammarco, Crenshaw, 1984; см. табл. 6.18, 6.19 и рис. 6.16, 6.17). Это и неудивительно, поскольку разные виды рифового зоопланктона, включая многие голопланктонные виды, выбирают в качестве дневного убежища разные типы донного субстрата (Emery, 1968; von Hentig, 1971).
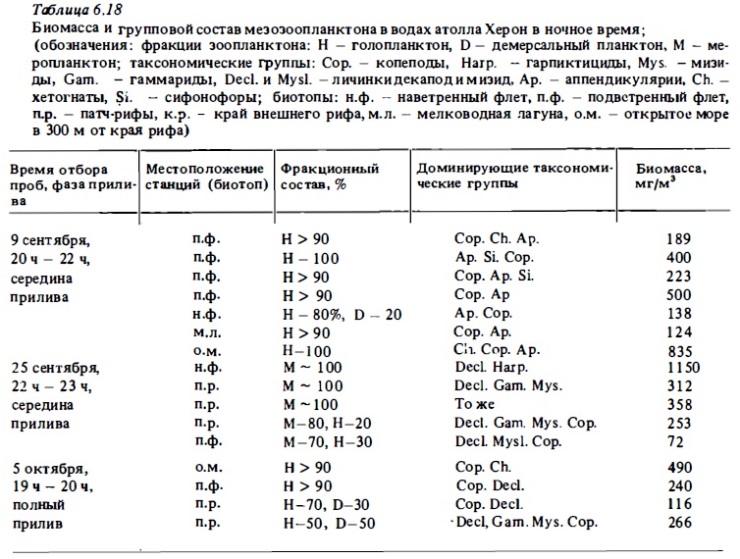
Среди голопланктонных форм зоопланктона рифовьк вод, так же как и среди зоопланктона открытого океана, доминирующими по численности обычно бывают каляноидные и циклопоидные копеподы и их личиночные стадии-науплиусы и копеподиты (см. табл.6.13, 6.15, 6.18, 6.19). Значительную численность дают и другие планктонные ракообразные: кладоцеры, амфиподы, мизвды, гарпактициды, остракоды (Renon, 1977). Многочисленными в составе голоинанктона рифа бывают хетогнаты, птероподы, а также аппендикулярии и их модель (см. табл.6.17, 6.18). В лагуне барьерного рифа у о-ва Новая Каледония копеподы играют подчиненную роль в составе зоопланктона (Champalbert, 1981). В составе голоинанктона лагун многочисленны кладоцеры и аппендикулярии. Многочисленны также гипонейстонные копеподы понтеллвды. Важным компонентом голопланктона рифов является микрозооинанктон: инфузории, зоовлагелляты, мелкие науплиусы копепод, но в рифовых водах до недавнего времени он практически не изучался (см.6.3.4). В табл.6.21 представлен список основных таксонов голопланктона вод коралловых рифов по результатам обработки сетных проб. Из табл.6.21 видно, что главное место по видовому разнообразию в составе голопланктона рифов занимают каляноидные копеподы. Весьма разнообразны также цихлоповдные копеподы, гарпактициды и более крупные представители планктонных раков-мизвды и амфиподы. При этом большая часть видов представленных в голопланктоне рифов входит в состав зоопланктона окружающих вод открытого океана (Goswami, 1983). Однако из этого не следует, что зоопланктон рифа является частью зоопланктона океана, который транспортируется ими на риф. Несмотря на присутствие большого числа океанских видов в его составе, зоопланктон рифа, как отмечалось выше, образует вполне обособленное сообщество, которое является частью экосистемы рифа и участвует в системе ее трофических отношений. Обособленность сообщества зоопланктона рифа, включая входящий в его состав голопланктон, следует уже из того, что численность тех же океанских видов в водах рифов в несколько раз, а иногда и на порядок величин и более выше, чем в окружающей риф пелагиали океана (см. табл.6.15). Из этого следует, что голопланктон вод рифа воспроизводится в пределах экосистемы рифа за счет ее энергетических ресурсов, а не заносится пассивно из океана. Наоборот, вода, уходящая с рифа в океан на его подветренной стороне, заметно обогащает зоопланктоном сами океанские воды (Johnson, 1949). В районах, где прибрежные воды богаты зоопланктоном, в ночное время с приливными течениями в зону рифа с окружающей акватории поступает в большом количестве голопланктон, который довольно быстро выедается донным сообществом рифа (см. табл.6.18) .
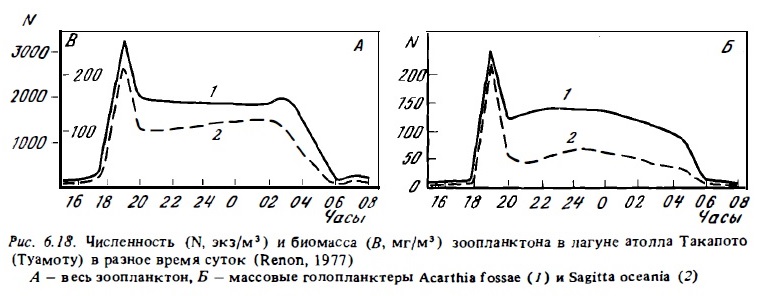
Существенно, что в условиях рифа океанские воды зоопланктона, например акарции, оитоны, начинают вести себя как другие компоненты типично рифового зоопланктона. В дневное время подавляющее большинство океанских видов голопланктона рифовых вод, как и весь остальной зоопланктон рифа, мигрирует ко дну, избегая выедания. При этом мигрируют не только быстро плавающие ракообразные и хетогнаты, но и медленно плавающие виды, такие, как аппендикулярии (см. табл.6.13, 6.14, рис.6.18). Многие представители голопланктона, включая океанские виды, ищут специфические убежища на дне и используют для этого неровности рельефа, колонии донных животных или образуют днем рои у дна (Emery, 1968; von Hentig, 1971; Hamner, Carleton, 1979).
Обособленные сообщества с преобладанием голопланктонных видов копепод могут образовываться даже в пределах одной и той же крупной лагуны в разных ее участках, отличающихся условиями среды. Так, путем математической обработки данных о составе зоопланктона в разных участках лагуны Б. барьерного рифа у г. Таунсвиль было доказано наличие в ней двух четко различимых сообществ зоопланктона. При этом локализация этих двух сообществ в пределах лагуны и их появление во времени в значительной степени зависят от объема речного стока, попадающего в лагуну (Sammarco, Crenshaw, 1984).
Как уже отмечалось выше, характерной особенностью сообществ рифового зоопланктона является присутствие в нем, а часто и преобладание по биомассе (и реже по численности) демерсальньк видов, которые появляются в составе зоопланктона рифа преимущественно ночью (см. табл.6.15—6.18). Днем они уходят в субстрат рыхлого грунта или на поверхность, а также прячутся в колониях горгонарий, в макрофитах и перифитонным обрастаниях скального грунта. Но главным их убежищем на рифе служат пористый известняк скальной поверхности флета и обломочный материал, который в большом количестве всегда присутствует в его донных биотопах (см. табл.6.20, рис. 6.17). Вследствие миграции со дна крупных демерсальных планктеров в толщу воды биомасса зоопланктона рифа возрастает ночью в десятки и сотни раз. Во много раз возрастает и его численность (см. табл.6.13, 6.14). В составе демерсального зоопланктона коралловых рифов доминирующее положение как по числу видов, так и по численности занимают различные ракообразные, и прежде всего копеподы, амфиподы и мизиды (Birkeland, Smalley, 1981). Многочисленными бывают и полихеты (табл.6.14, 6.16).
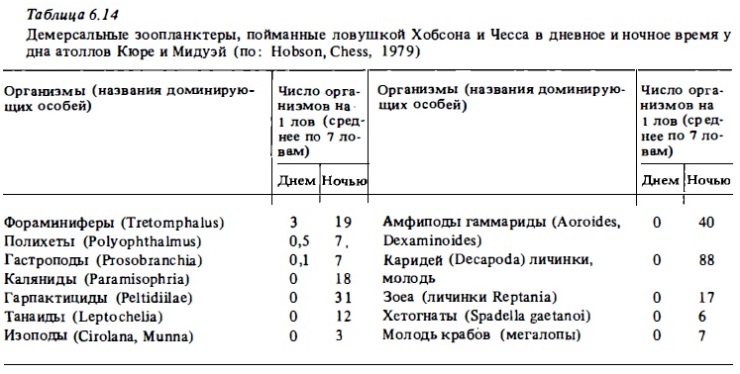
Значительную долю ночного, а также и дневного зоопланктона коралловых рифов составляют меропланктонные организмы, представленные главным образом личиночными стадиями и молодью ракообразный, моллюсков и полихет, а также личинками иглокожих, асцидий, кишечнополостным (кораллов, гидроидов). К меропланктону рифа относят и личинок рыб на ранних стадиях развития, а также икринки. Как видно из данным о суточным изменениях в составе зоопланктона рифов (табл.6.14), меропланктон, как и демерсальный зоопланктон, днем прячется в убежищах на дне и выходит в толщу воды преимущественно в ночное время, хотя некоторые его представители, такие, как, например, личинки моллюсков велигеры, часто в большом количестве встречаются и днем в планктоне рифа (см. табл.6.15, 6.17) . В табл.6.22 представлен список основным таксонов беспозвоночных, формирующих демерсальный зоопланктон и меропланктон вод коралловым рифов. Как уже отмечалось, доминирование тех или иных групп этих планктеров в пробах зоопланктона и сам его видовой состав находятся в большой зависимости от применявшихся орудий лова, от времени и места сборов (Alldredge., King, 1977, 1980; Hobson, Chess, 1979; Мс William et al., 1981). Поэтому у разных авторов заключения на этот счет весьма неоднозначны. Особенно это касается значения копепод как компонента демерсального зоопланктона, мигрирующего ночью в толщу воды из донного субстрата.
Заключения таких авторов, как Портер и Портер (Porter, Porter, 1977), Портер и др. (Porter et al., 1977), Олдридж и Кинг (Alldredge, King, 1977), о значительном преобладании копепод по численности над всеми другими видами демерсального зоопланктона, попадающего в ловушки, установленные на дне, явно некорректны (Hobson, Chess, 1979). Последние авторы убедительно показали, что ловушки, которые упомянутые ранее авторы использовали для учета зоопланктона, поднимающегося со дна, прилавливали в большом количестве со стороны голопланктонные копеподы и мизиды. В уловах демерсального зоопланктона, полученных с помощью усовершенствованным ловушек, конструкция которых снижала (но, по-видимому, не исключала полностью) возможность прилова, процент копепод составлял не 80-90, а 55-60% (см. рис. 6.16; Мс William et al., 1981; Birkeland, Smalley, 1981).
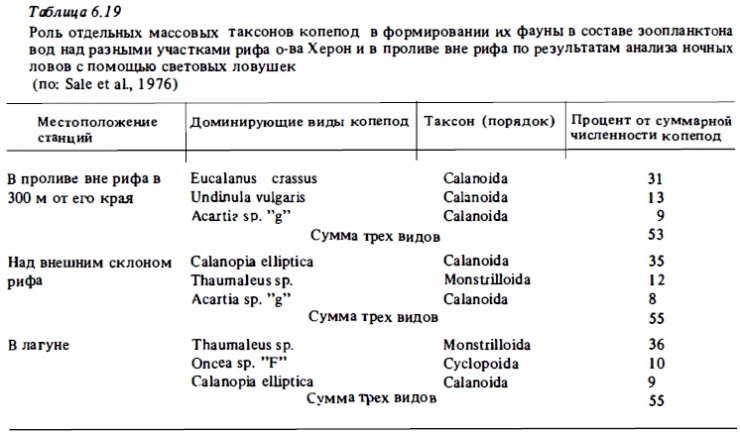
Учет зоопланктона рифа в ночных уловах световых ловушек (Sale et al., 1976) показал четкие различия в таксономическом составе копепод, обитающих в зоне рифа и в прилегающих водах открытого океана. В уловах, сделанных над патч-рифом в лагуне, доминировали не каляниды, как это обычно бывает в открытом море, а циклопиды и монстриллиды (Thaumallus sp.). Последние в водах открытого моря встречаются крайне редко. Даже над внешним склоном рифа, где должно происходить интенсивное смешение фаун зоопланктона рифа и открытого моря, доминировали виды, живущие в рифовых водах (Calanopia elliptica Thaumaleus sp.), а не обычные для вод открытого океана Eucalanus crassus и Undinula vulgaris. В пробах из ловушек, полученный на флете барьерного рифа у о-ва Муреа в зарослях макрофитов, преобладали каляниды и гарпактициды. В аналогичных пробах из биотопов лагуны атолла Такапото доминировали амфиподы и танаиды (Birkeland, Smalley, 1981).
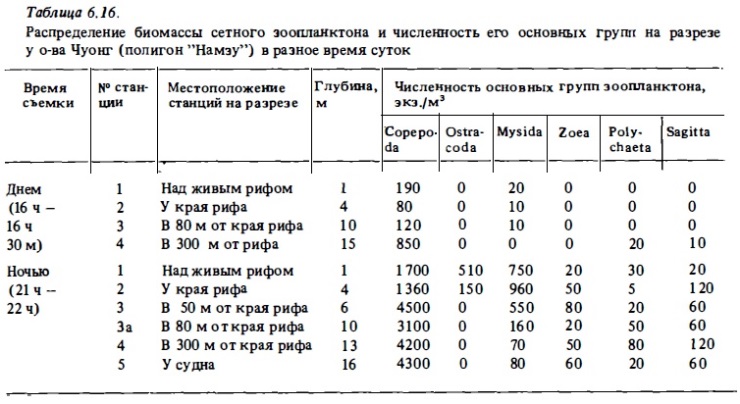
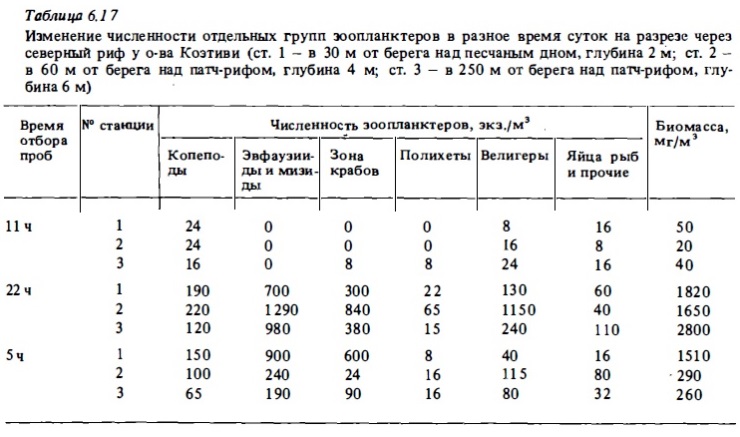
В наших ночных уловах планктона в водах над рифами в одном случае (риф у островов Намзу) доминировали копеподы, численность которых достигала очень высоких величин — до 4500 экз/м3 (см. табл.6.16) . На рифе у о-ва Коэтиви в составе ночного зоопланктона доминировали крупные меропланктонные и голопланктонные виды ракообразный (мизвды, амфиподы, эвфауэииды (см. табл.6.17). В первом случае (у островов Намэу) протяженность окаймляющих рифов бьша наибольшей — около 80 м. В этих условиях следовало ожидать значительного влияния богатого копеподного зоопланктона прилегающих вод Сиамского залива на состав зоопланктона рифов. Однако даже в этих условиях, несмотря на преобладание копепод на всех станциях ночного разреза у о-ва Чуонг, основную биомассу формировали рифовые виды меропланктонных и гологшанктонных миэвд и амфипод, численность которых у края рифа была близка к численности копепод при общей биомассе рифовых видов, более чем на порядок превышавшей биомассу последних.