
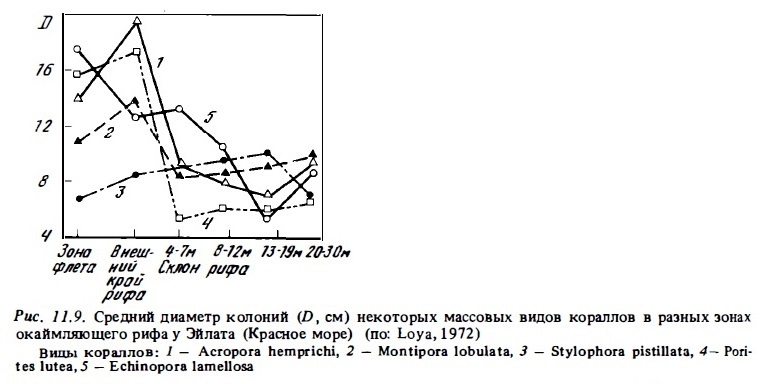
В отношении размерного состава кораллов в сообществах рифов, то в них, как правило, преобладают среднеразмерные молодые колонии диаметром 5—15 см (рис. 11.9) и площадью от 200 до 600 см2 (Loya, 1972; Sheppard, 1980). В составе видовых популяций у многих видов кораллов (Acropora, Galaxea, Echinopora) по наибольшему числу колоний выделяются две размерные группы: очень молодае кораллы с малой площадью колоний (20-200 см2) и кораллы средних размеров с площадью колоний от 4 до 20 дм2 (рис. 11.10). У других видов первая размерная группа моло-дах малоразмерных колоний немногочисленна, и в составе их популяции преобладают кораллы средней размерной группы (Porites, Lobophyllia, Cteiiella, Stylopora). В разных зонах рифа размерный состав видовых популяций кораллов может меняться (Loya, 1972). При этом в пределах отдельных видов средние размеры их колоний бывают выше в верхней прибойной зоне склона и на неосушаемом флете, чем на глубинах 5-10 м. Вниз по склону, глубже 10-20 м, средний размер колоний большинства кораллов и их возраст увеличиваются. Так, на рифах Ямайки на глубинах 10-20 м колонии площадью более 200 см2 составляли 9%, а на глубине 35 м — 17%. В мелководной зоне смертности кораллов и число оседающих личинок выше, чем на нижних горизонтах склона рифа. Поэтому в верхних зонах оборачиваемость видовых популяций кораллов происходит интенсивнее, чем на нижних горизонтах, где слабее физический стресс (Huges, 1982) . Средний возраст колоний кораллов в сообществах внешнего склона оценивается для быстрорастущих ветвистых кораллов-акропорид и псаммокоры в 2-5 лет и для медленно растущих поритид и фавиид — 7—11 лет. В составе популяций некоторых массивных форм кораллов, таких, как Montastrea пли Porites, часто преобладают крупные старые колонии диаметром 1-2 м, возраст которых достигает десятков и сотен лет (Isdale, 1984; Cameron, Endean, 1985).
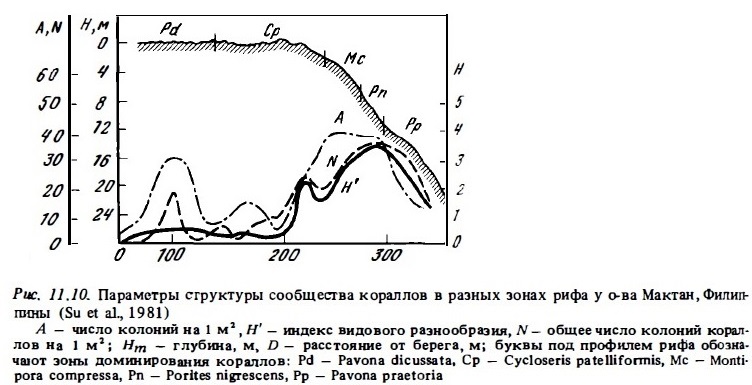
При среднем покрытии кораллами поверхности дна в сообществах внешнего склона рифа в пределах 40—80% среднее число колоний кораллов в расчете на 1 м2 варьирует обычно в пределах 20-40 экз. (рис. 11.11). На флете оно снижается до 5-20 экз./м2. Максимум численности колоний на 1 м2 дна до 60 экз. — обнаруживается в верхних горизонтах склона и в зоне опор. При этом преобладающая их часть обычно представлена 2—4 доминирующими видами твердых или мягких кораллов. Эти же вида, как правило, занимают более половины всей площади дна, покрытой кораллами (см. табл. 11.4, 11.7). На долю одного доминирующего вида может приходиться до 10-20% всего числа колоний кораллов в сообществах. На многих рифах значительная, а иногда и преобладающая часть общего числа колоний представлена мягкими (восьмилучевыми) кораллами. В частности, на карибских рифах они могут представлять до 80-90% всей численности колоний в биотопе (Jaap, 1985).
Как уже отмечалось выше, общее число видов кораллов и показатели их видового разнообразия, относительно низкие в лагуне и на флете, резко возрастают в сообществах верхних горизонтов внешнего склона — до глубин 20—30 м (Loya, 1972; Sheppard, 1980; Syet al., 1981; Latypov, 1985; Титлянов и др., 1988). На глубинах от 5 до 30 м на рифах Индо-Тихоокеанского региона число видов в зонах внешнего склона составляет от 30 до 100 и на рифах Западной Атлантики — 15-20 видов (см. рис. 11.4-11.7) . В соответствии с этими графиками общее число видов кораллов в сообществах склона является в определённой степени функцией глубины, нарастая до глубин 20-40 м. Зональность в структуре сообществ проявляется в составе доминирующих в них видов кораллов, в преобладании тех или иных ростовых форм их колоний (рис. 11.12), а также в появлении в отдельных зонах характерных для них высокоспециализированных видов. Однако в большинстве биотопов рифа (исключая дно) преобладающее место в составе сообществ занимают оппортунистические виды кораллов. Их характерная особенность состоит в способности к широкой фенотипической изменчивости. Последняя выражается в их способности образовывать в зависимости от конкретных условий среды адаптивные ростовые формы колоний (Abe, 1937; Boschma, 1956; Goreau, 1963; Stoddart, 1969; Rosen, 1971; Hubbard, 1973; Foster, 1979; Преображенский, Иванов, 1982; Brakel, 1983). Главными факторами среды, которые определяют характер этой адаптативной изменчивости формы колоний кораллов, являются волновой стресс, освещенность и занос осадками (Pichon, 1974; Dustan, 1975; Wainwright, Koehl, 1976) . Общая тенденция адаптивных изменений формы кораллов, имеющих ветвистые или столбчатые колонии, под влиянием волнобоя состоит в образовании более мощных укороченным отростков. Таковы акропоры А. palifera, А. palmata, А. digitifera, Pocillopora verrucosa. Массивные и листовидные кораллы (Porites, Goniastrea, Goniopora, Galaxea, Pavona, Montipora) в зоне волнобоя образуют уплощенные или корковидные колонии (Morton, 1974; Brakel, 1983). Тенденцш адаптивных изменений колоний кораллов с глубиной под влиянием убывания освещенности и снижения гидродинамического стресса выражается в уплощении колоний в образовании корковидных форм. Такие формы колоний в глубинных зонах рифа образуют многие виды ветвистых и массивных кораллов: Acropora, Stylophon (S. mammilata), Pavona, Porites (Р. asteroides), Montastrea(M. annularis), Favia, Psammocora, Echinopora (Kawagati, 1937; Wells, 1957; Lewis, 1960; Goreau, 1963; Faure, 1974; Veron, 1981).
В зонах слабого и умеренного гидродинамического стресса на подветренных и лагунных рифах, в средней части внешнего склона обычно отмечается наибольшее разнообразие видового состава и форм колоний (Connell, 1978; Grigg, 1981). Отбор доминирующих видов идет в этих условиях по многим факторам, в числе которых важное место приобретает конкурентоспособность, устойчивость к заилению и выеданию, способность к использованию света, к гетеротрофному питанию зоопланктоном.

Несмотря на способность большинства герматипных склерактиниевых кораллов существовать во всем диапазоне глубин рифа - от его верхних горизонтов до подножия, в каждой отдельной зоне рифа в составе их сообществ часто доминируют один или несколько видов. При этом доминирование наиболее четко бывает выражено в его мелководных зонах, которые в значительной степени подвержены воздействию физического стресса: волнению, осушке, заносу осадками, колебаниям температуры и солености. На рифах Индо-Тихоокеанского региона в число таких руководящих видов прибойной зоны входит ряд родов: Pocillopora, Porites и Millepora, а также акропоры: А. hyacinthus, А. cuneata, А. humilis, А. digitifera, А. leptocyathus, А. hebes, А. fornose, А. pulchra, А. palifera, Stylophora pistillata, Echinopora gemmaceae. На рифах Атлантики в числе таких видов: А. cervicornis, А. palmata, А. prolifera, Millepora alcicoruis, Porites furcata. Роль руководящих видов кораллов с массивными или корковидными колониями в прибойной зоне индо-тихоокеанских рифов выполняют фавииды: Goniastrea, Favia, Favites, Montipora, Galaxea, Pavona, Psammocora, Platygyra, а на рифах Атлантики — Diploria, Montastrea, annularis, Porites asteroid. Некоторые из этих видов хорошо переносят наряду с волнобоем также и другие виды физического стресса. На осушаемом флете с доминированием перечисленных видов кораллы выживают при их полной осушке, в сумме от 10 до 80 часов за месяц при продолжительности отдельных периодов осушки в течение фазы отлива от 1 до 4 часов (Ditlev, 1982). Особенности стойкой к осушке оказалась А. pulchra, которая часто преобладает в зоне осушаемого флета. Она выдерживает полную осушку в течение 2-4 ч и не погибает при этом под дождем и под солнцем (Vaughan, 1916; Ditlev, 1978). Кораллы Pocillopora и Montipora хорошо переносят также типичные для мелководных зон рифов стрессовые воздействия, как мутная вода и занос осадками, высокая температура и распреснение (Loya, 1972), соленость до 40°/оо (Wells, 1957; Smith, Jokiel, 1978). Однако, если в условиях интенсивного заиления в донных осадках начинается бактериальная редукция сульфатов до сероводорода, кораллы быстро погибают (Ditlev, 1978).
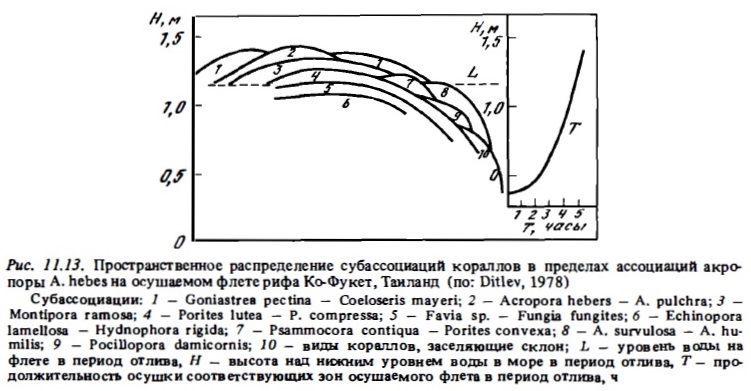
На многих приподнятых индо-тихоокеанских рифах, рано достигших уровня моря, самая высокая зона их осушаемого флета обычно достраивается кораллинами. Зона роста кораллов начинается на нем лишь вблизи прибойного края рифа (Rosen, 1971a). В этих сообществах доминируют при сильном прибое Acropora pulchra и Porites lutea. Если прибой слабее, в сообществах доминирует ассоциация Montipora — Porites — Goniastrea retiformis. В условиях большой мутности вод на осушном флете хорошо выживает и быстро восстанавливается после катастрофических воздействий ассоциация: Porites lutea — Acropora hebes, а при малой мутности вод — ассоциация Stylopora pistillata — MiUepora dichotoma — Porites lutea — Pocillopora damicornis и А. digitifera — А. cuneata - Montipora foliosa (Pichon, 1964; Taylor, 1968; Loya, 1976а, Bothwell, 1984) (рис. 11.13).

На погруженном флете при умеренном или малом волнобое на подветренных рифах или на рифах, защищенных барьерами, видовое разнообразие кораллов бывает достаточно высоким. Примером может быть неосыхающий флет атолла Херон, где на площади 120 м2 было зарегистрировано 40 видов герматипных кораллов при покрытии кораллами менее 10% поверхности флета. При этом около половины всех видов были представлены массивными формами (Cameron, Endean, 1985). На многих рифах наибольшее число видов обнаруживается именно в сообществах неосущаемого флета. В их составе, как правило, доминируют массивные столбчатые или листовидные кораллы родов: Porites, Goniastrea, Goniopora, Galaxea, Favia, Montastrea, Montipora. Ветвистые кораллы представлены эдесь родами: Acropora, Pocillopora, Seriatopora, Millepora.
В приглубых биотопах внешнего склона рифа, где число видов и показатели видового разнообразия кораллов достигают максимума, доминирование отдельных видов в сообществах выражено слабо. Наибольшего развития в этой зоне достигают виды, имеющие массивные столбчатые или листовидные колонии. На индо-тихоокеанских рифах они представлены родами: Porites, Pectinia, Symphillia, Coscinarea, Merulina, Montipora, Goniastrea, Montastrea, Pashyseris, Hydnophora, Echinopora (Done, 1983; Sheppard, 1980, 1982), и на рифах Атлантики — родами: Montastrea, Siderastrea, Agaricea, Millepora, Dichocoenia (Kuhlmann, 1974, 1975; Abel et аl., 1983). Встречаются в этой зоне ветвистые оппортунистические кораллы Pocillopora, Stylophora, Seriatopora и несколько видов акропор.
В сообществах кораллов, которые обитают в нижней части склона у подножия рифа, видовое разнообразие сохраняется обычно на высоком уровне (см. рис. 11.3-11.5), причем в их составе преобладают герматипные виды, несмотря на то что их развитие здесь угнетается недостатком света (ФАРп 5%). В нижней части склона при таком уровне освещенности в среднем около 90% видов представлены герматипными склерактиниями, способными к световой адаптации, и всего около 8-10% видов представлены олигатными герматипными и агерматипными видами, которые обитают только в глубинной зоне. В составе первой, более многочисленной группы кораллов, заселяющих глубинные зоны индо-тихоокеанских рифов, - представители родов: Stylophora, Porites, (Р. australiensis), Leptastrea, Asteropora, Pavona, Echinopora, Fungia (F. granulosa) Pocillolopara (Р. solida, Р. verrucosa), Synarea, Stylophora (S. kiihlmani), Podabacia, Coscinarea, Mycedium. Колонии этих кораллов приобретают в глубинной зоне уплощенную форму, более тонкую архитектонику, адаптивно более приспособленную к улавливанию слабого света (Bouchon, 1981; Fricke, Schuhmacher, 1983; Титлянов и др., 1988).
На рифах Атлантики состав сообществ кораллов в нижней части склона становится сходным с таковым в Индо-Пацифик'е. В их составе: Leptoseris, Agaricia, Scolymia, Madracis formosa, Montastrea cavernosa. Подчеркивая видимое исчезновение разницы в видовом составе сообществ кораллов в глубинных зонах фор-рифа этих ареалов, Д. Кюльман (Kuhlmann, 1983) называет их глубинными ассоциациями. В составе таких ассоциаций преобладают уплощенные или листовидные формы колоний кораллов, которые в мелководных зонах могут иметь ветвистую или глобулярную массивную форму (Schuhmacher et al., 1985). В этих ассоциациях значительную роль играют также агерматинные кораллы: склерактинии, антипатарии, горгонарии и альционарии.
В составе облигатных глубоководных герматипных склерактиний, заселяющих подножие внешнего склона рифа, преобладают высокоспециализированннге виды рода Leptoseris (L. mycetoserioides, L. explanata, L. porosa), имеющие специальную световоспринимающую систему (Fricke, Schuhmacher, 1983) и способные выживать при освещенности ниже 1% ФАРп. К таким специализированным кораллам относятся также Cycloseris cyclolithes, С. patellifonnis, Pachyseris rugosa, Р. speciosa, Scolymia vitiensis, Leptastrea bewickensis. Фауна агерматипных склерактиний у подножия рифа достаточно богата и представлена видами родов: Dendrophyllia, Tubastrea, Astrangia (А. solitaria), Phyllangia (Р. americana).
При изучении сообществ кораллов у подножия внешнего склона рифов до глубин 200 м интересные результаты были получены с помощью подводных аппаратов (Lang, 1974; Fricke, Schuhmacher, 1983; Fricke, Hottinget, 1983; Reed, 1985; Fricke, Meischner, 1985). Герматипные кораллы Agaricia grahamae и Montastrea cavernosa были обнаружены на глубинах до 113 м, где освещенность бьша около 0,15% ФАРп, а коралл Eusmilia fastigata — на глубине 140 м. На склонах тихоокеанских атоллов некоторые герматип-ные кораллы бьши найдены на глубинах более 140 м, в том числе Pachyseris fastigata — на глубине 200 м, Coscinarea monilis — на глубине 270 м, Psammocora explanulata, Scolymya lacera, Leptoseris porosa и L. fragilis на глубинах 140-165 м (Kuhlmann, 1983). Эти глубины намного превьшают глубину положения компенсационной точки фотосинтеза, которая даже у самых адпатированных к недостатку света кораллов находится в прозрачных океанских водах на глубинах не более 100 м (Fricke, Schuhmacher, 1983). В составе агерматипных кораллов на глубинах более 100 м были обнаружены Madracis interjecta, М. myriaster, Dendrophyllia minuscula, Oculina varicosa. Наблюдения показали, что ассоциации кораллов, которые заселяют подножие рифа на глубинах более 80—100 м, не образуют трехмерных рифовых построек. Ввиду замедленной скорости кальцификации и уплощенности колоний кораллов они образуют на этих глубинах отдельные фации или биогермы, пятнами разбросанные по склону. Интенсивность кальцификации здесь не превышает скорости биоэроэии известкового материала. Поэтому карбонатный материал, который накапливается этими сообществами, разрушается и стекает по склону, не образуя рифа (Fricke, Hottinger, 1983; Schuhmacher, 1985). Аналогичные постройки типа биогерм образуют глубоководные склерактиниевые кораллы с известковым скелетом. В их чиспе: Lophelia, Solenosmylia, Madrepora oculata, Goniocorella, DesniophyUum, Oculina varicosa. Эти виды образуют биогермы на континентальном склоне до глубин более 1000 м (Келлер, 1976; Zibrovius, 1980; Cairns, Stanley, 1981).
Своеобразные сообщества кораллов заселяют донные биотопы рифов, занятые рыхлыми осадками: илами, эаиленными коралловыми песками и частыми коралловыми песками. Поселение и нормальный рост прикрепленных форм кораллов здесь затруднены. Тем не менее биотопы мягкого дна оказываются часто достаточно плотно заселенными кораллами, среди которых преобладают свободно живущие вины склерактиний, представленные одиночными полипами. В их числе: Fungia, Diaseris и Cycloseris из сем. Fungiidae, Heteropsammia из сем. Dendrophylliidae и Heterocyathus из сем. Caryophilliidae (Goreau, Yonge, 1968; Pichon, 1978; Fisk, 1983; Latypov, 1985). На мягком дне встречаются и некоторые колониальные кораллы. Ветвистые кораллы Acropora, Pcillopora, Stylophora селятся на обломках крупных колоний кораллов, на раковинах тридакн. Два последних их рода, а также некоторые другие массивные и пластинчатые кораллы, такие, как Goniopora и Euphillia, Povona, Agaricella, Psammocora, заселяют биотопы дна, образуя небольшие округлые колонии, свободно лежащие на грунте.
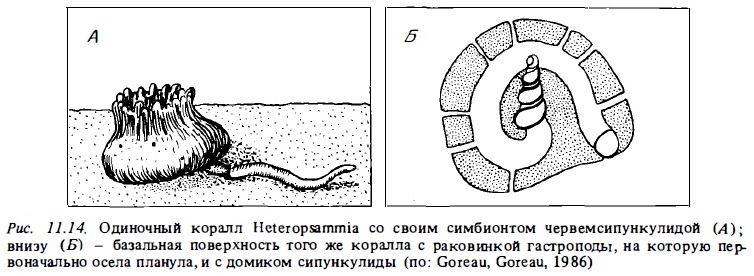
Основным экологическим фактором, угнетающим рост кораллов в этих биотопах, является занос их осадками или погружение в рыхлый осадок под собственной тяжестью. Разные их вины используют разные механизмы для преодоления заноса колоний осадком. Ветвистые кораллы, прикрепленные к крупным обломкам или раковинам, обладают высокой скоростью роста и избегают заиления, сидя на своеобразном "корне" с подошвой, погруженной в осадок. Последний представляет собой отмершее оснозание колонии, прикрепленное к обломку. Округлые свободные колонии избегают заноса осадками за счет того, что их постоянно двигают донные животные: рыбы и гастроподы, которые во время кормежки ищут под ними корм (Glynn, 1974). Фунгии обычно заселяют участки дна, расположенные в активных гидродинамических зонах: на флете, в каналах и каньонах. Они передвигаются по дну, занятому песком или обломочным материалом, под действием волн, избегая таким путем заноса осадком (Stoddart, 1969). Они способны интенсивно сбрасывать с себя оседающую взвесь (Schuhmacher, 1977). Одиночные свободно живущие полипы фунгид — Helofungia, Diaseris, Cycloseris — могут выбираться из толщи осадка и даже немного передвигаться, упираясь в грунт своими длинными щупальцами (Gill, Coates, 1977). И все же наиболее специализированными свободно живущими кораллами мягкого дна являются герма-типные одиночные кораллы Heteropsammia и Heterocyathus. Они не встречаются в других биотопах рифа. Дпя них характернен очень совершенный механизм сбрасывания оседающей на них взвеси, основанный на взаимодействии движений щупалец и ресничного аппарата (Fisk, 1981). Однако наиболее удивительная их способность состоит в том, что они могут передвигаться по поверхности грунта аналогично актиниям, которых "возят" на себе крабы-опиельники. Таким "извозчиком" служит для них живущий с ними в сожительстве червь-сипуккулида Aspidosiphon. Этот червь-сибионт "возит" коралл по поверхности заиленных коралловых песков со скоростью 0,5-1 м за ночь (рис. 11.14).
Плотность сообществ кораллов мягкого дна может быть очень высокой. Массовые их вид — Heteropsammia, Diaseria, Cydoseris — образуют поселения плотностью до 1000 экз./м2 при размерах отдельных особей-кораллитов 0,5—2,5 см (Latypov, 1985). Высокую плотность можно встретить и в поселениях фунгий на коралловых песках: 3-5 особей размером 5-15 см на 1 м2.