
Строение полипов склерактиниевых кораллов в своей основе сходно со строением актиний. Они имеют бокалообразное тело и шесть или кратное шести количество простых гладких щупалец. В коечной полости полипа имеется шесть или кратное шести количество мезентериальных септ-перегородок. От их краев отходят мезентериальные нити, которые гистологически отличны от тканей эндотелия, выстилающих кишечную полость полипа. Мезентериальные несут в себе группы клеток железистого типа и являются органами пищеварения полипа (Yonge, 1968). У большинства склерактиниевых кораллов их длина такова, что позволяет им выходить наружу через рот или через стенку кишечной полости полипа и переваривать вне тела пойманные полипом крупные жертвы, которые он не может проглотить.
Отличие полипов склерактиний от полипов актиний состоит в их способности строить наружный известковый скелет (Рифы, 1987). Скелет индивидуального полипа, именуемый кораллитом, состоит из базальной пластинки, которая окружена бокаловидной текой. От теки внутрь ее отходят радиальные перегородки-септы.Размеры полипов у разных групп герматидных склерактиний варьируют от 1 мм до 30—40 см. Размеры колоний разных систематических групп склерактиниевых кораллов также варьируют в очень широких пределах: от нескольких сантиметров (Psammoco, Galaxea) до 2-3 м. Средние же их размеры в сообществах риф-флета и склонов обычно бывают небольшими (10-20 см), так же как и их средний возраст (3-6 лет), поскольку смертность кораллов с возрастом резко возрастает (Bak, Luckhurst, 1980).
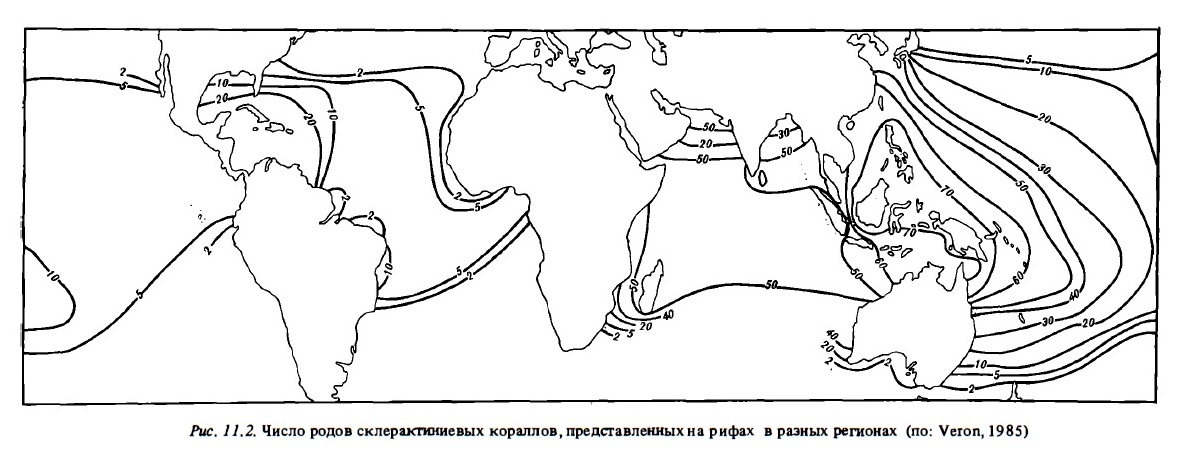
Сообщества герматипных склерактиниевых кораллов, населяющие коралловые рифы, отличаются большим видовым разнообразием (Potts, 1983). Общее число истинных их видов достигает 530 при общем числе их родов около 110. Они обитают преимущественно лишь в пределах тропического шельфа в верхних его горизонтах до глубин 80-100 м, где температура вода не опускается ниже 18-19°С. Число родов агерматипных склерактиниевых кораллов еще выше, чем склерактиниевых. Всего их около 150 родов. Они заселяют донные биотопы всех зон Мирового океана до полярных широт и до глубин более 1500 м (Келлер, 1976; Cairns, 1982). Видовое разнообразие фауны склерактиниевых герматипных кораллов, равно как и ее состав, варьируют на рифах разных регионов Мирового океана. Общее число родов этих рифостроящих кораллов достигает максимума на рифах в районе Индонезни-Филиппин-севера Австралии, где можно насчитать до 70 родов и 250-350 видов герматипных склерактиний (Porter, 1972; Veron, 1985). Схема распределения численности их родов в разных регионах представлена на рис. 11.2. Беднее всего по числу родов этих кораллов рифы Атлантики — всего около 20.
Такая неравномерность распределения фауны наиболее поздних по своему происхождению склерактиниевых кораллов в бассейнах Мирового океана является свидетельством ее относительной молодости. Положение о том, что формирование фауны склерактиний на современных рифах продолжается интенсивно и в наше время (Potts, 1981; Veron, 1985), подтверждается также и фактом резкой обособленности их фаун в Атлантике и в Индо-Тихоокеанском регионе. В Атлантическом океане, более молодом по возрасту, фауна склерактиний в 3-4 раза беднее, чем в Индо-Тихоокеанском бассейне. Только 6 родов из 60-70 родов, обитающих на индо-тихоокеанских рифах можно встретить и на рифах Атлантики. В их числе Acropora, Cladocora, Favia, Madracis, Porites и Siderastrea. При этом на рифах Атлантики практически все вида указанных общих родов иные, чем в Индо-Пацифике. Общим для рифов этих регионов Мирового океана можно считать лишь один вид рода Siderastrea (S. radians). Среди 150 агерматипных склерактиний общими для обоих бассейнов являются около 20 родов и всего
3-5 видов, в числе которых такие массовые их виды, как Tubastrea aurea и Magracis asperula. Сама по себе обособленность фауны даже агерматипных склерактиний Атлантического и Индо-Тихоокеанского регионов также весьма показательно демонстрирует общую относительную молодость фауны склерантиниевых кораллов Мирового океана. Действительно, фауна агерматипньк склерактиний, обитающая в пределах умеренных и высоких широт, в меньшей степени изолировала географически, имея канал связи между этими двумя регионами в пределах Южного океана.
Как уже отмечалось ранее, наибольшее видовое разнообразие фауны герматипных склерактиний зарегистрировано в рифовых комплексах на стыке Индийского и Тихого океанов в районе Индонезии, Филиппин, и северной части Большого барьерного рифа. Число их родов составляет здесь 60-70 и число видов 250-350. Достаточно богата фауна кораллов на рифах центральной части Индийского океана, в Красном море и в западной части Тихого океана: 50-60 родов и 150-250 видов (см. рис. 11.2). На рифах Западной Атлантики обычно встречается 15-25 родов герматипных склерактиний и 30-60 их видов. Самая бедная фауна склерактиний характерна для восточных побережий Африки и центральной Америки: 3-5 родов и 7-12видов. На отдельных рифах в центре Индо-Тихоокеанского региона можно найти до 30-50 родов и 80-200 видов герматипных склерактиний. На отдельных рифах Западной Атлантики обнаруживают обычно 10-20 родов и 15-30 их видов.
Высокий уровень видового разнообразия современной фауны склерактиний непосредственно обусловлен многократным прерыванием процесса их видообразования, вызванным частыми значительными колебаниями уровня океана в плиоцене. Процесс видообразования и дифференциации фауны современных герматипных склерактиниевых кораллов начался сравнительно недавно, всего 20-30 мнл лет назад в мооцене после закрытия Тетиса и начавшегося образования Атлантического океана. До этого периода фауна герматипных склерактиний, которая появилась в мезозое около 240-600 мнл лет назад, была достаточно равномерно распределена в мелководных зонах эпиконтинентальных морей мезозойского Тетиса и занимала в их донных биотопах одно из ведущих мест. Большинство современных их семейств и родов сформировались уже к середине четвертичного периода. В позднечетвертичном периоде произошло разделение фауны склерактиний на Индо-Тихоокеанскую и Атлантическую, которое значительно ускорилось после образования 3—5 млн лет назад Панамского перешейка, полностью разделившего тропическую зону обоих бассейнов (МсСоу, Heck, 1976). Затем в периоды оледнения, которые в плиоцене, а затем и в плейстоцене следовали одни за другим, начались сильные колебания уровня океана, вызывавшие периодические ослики и гибель фауны герматипных склерактиний как в Атлантике, так и в Индо-Тихоокеанском регионе. Всего только шесть родов герматипных склерактиний, живших до закрытия Панамского перешейка в обоих бассейнах, пережили оледнения и осушки и сейчас встречаются по обе его стороны в Атлантике и в Индо-Пицифике. Эти роды были перечислены выше. Однако, даже и в пределах этих родов составляющие их вида в обоих бассейнах иные, образовавшиеся уже в плейстоцене (Stehli, Wells, 1971; Stanley, 1981). От этих осушек и похолоданий, связанных с оледнениями, погибали прежде всего именно герматишые склерактниии, которые живут только в верхних горизонтах тропического шельфа. В периода между оледенениями, когда остатки пережившей их фауны начинали заново развиваться и строить новые рифы, начиналось интенсивное видообразование (Potts, 1981), о чем и свидетельствует полная разделенность фауны Атлантики и Индо-Пацифики на уровне видов. Современные рифы, как мы знаем, очень молода. Им всего 7-10 тыс лет. Их рост и соответственно формирование фауны рифострояших кораллов продолжаются и в наше время, что не в последнюю очередь объясняет высокое их видовое разнообразие в пределах отдельных рифов (Macintyre, Glynn, 1976; Potts, 1981; Randall, 1985).
Исследователи, изучавшие состав фауны центральной части Индо-Тихоокеанского региона, восточной части Индийского океана и Красного моря, обратили внимание на поразительную гомогенность фауны в пределах отдельных рифов этих районов, разнесение на большие расстоянии. Эти сведения были интерпретированы как свидетельство в пользу того, что герматидные склерактнии Индо-Тихоокеанского региона пройти длительное и непрерывное эволюционное развитие в биогсографическом центре своего происхождения, где они сохранялись в периоды оледенения и откуда постепенно распространялись по всему бассейну (Newell, 1971; Rosen, 1971; 1984; Briggs, 1974). Однако такую точку зрения в настоящее время принять трудно, имея надежные доказательства прерывистости эволюции кораллов и роста самих рифов в конце третичного-четвертичном периоде (МсСоу, Heck, 1976; Potts, 1981; Veron, 1985). Согласно последним авторам, фауна герматипных склерактиний лучше всего перевивала периоды оледенений и падения уровня океана в мелководных бассейнах в зоне Филиппни-Индонезии. Поэтому данный регион, характеризующийся в настоящее время наибольшим разнообразием фауны рифообразующих склерактиний (Pichon, 1977) следует считать не центром про^ого их видообразования, а скорее центром их сохранения в периоды падения уровня океана. Отсюда при последующем подъеме уровня океана кораллы, имеющие пелагические личинки-планулы, могли сравнительно быстро распространяться вдоль побережий и между островами и вскоре равномерно вновь расселялись по всей зоне континентального шельфа Индийского и западной части Тихого океана и их островов, где сосредоточено более 80% всех коралловых рифов Мирового океана. Как показали эксперименты, плануда кораллов могут недели и даже месяцы жить в аквариумах не оседая, что доказывает возможность их разноса на большие расстояния.
Зоогеографическая широтная граница распространения герматипных склерактиний-рифостроителей определяется нижним пределом температуры, ограничивающим их рост (18-19°С). Причина существования такой температурной граней их развития скорее всего связана с тем, что при снижении температуры замедляется процесс кальцификации (Crossland et al., 1984). Для поддержания его на высоком уровне, достаточном для формирования рифов при пониженной температуре, требуется больше света. Таким образом, световой порог процесса кальцификации у кораллов в умеренных и холодных водах должен быть выше, в то время как в высоких широтах освещенность в целом падает. Поэтому склерактинии в этих условиях не могут достичь скорости кальцификации, достаточной для компенсации и превышения скорости эрозии рифа, что является непременным условием его роста и соответственно формирования полноценных сообществ кораллов-рифостроителей.