
В своей работе о механизме захвата и заглатывания пищи полипами склерактиний Янг (Yong, 1930) придает большое значение взаимодействию щупалец с деятельностью ресничного аппарата, который, по его мнению, обеспечивает транспорт ко рту и заглатывание пойманных и обездвиженных жертв. Последующие исследования показали явную ошибочность ряда его заключений, связанных с описанием роли ресничного аппарата в захвате пищи у склерактиний (Lewis, Price, 1975). Ошибка Янга состояла не в преуменьшении этой роли, а в неверном понимании самого механизма участия ресничного аппарата в захвате и заглатывании пищи кораллами. Дело в том, что ресничный аппарат полипа может участвовать в процессе захвата пищи одновременно с выполнением своей основной функции — очисткой полипа от взвеси. Для этого ему не надо менять направление биения ресничек, поскольку реснички как таковые фактически не играют главной роли в транспорте пищи ко рту и обращать направление их блиения не требуется, так как пища захватывается и переносится ко рту главным образом слизевыми тяжами, которые втягиваются полипом в рот и заглатываются. В этом случае перенос пищевых частиц будет происходить в направлении, противоположном биению ресничек. Обращение же биения ресничек у фунгий и лептастрий, описанное Янгом, а до него и другими авторами, равно как и заключение о постоянном биении ресничек ко рту у агирицид, можно рассматривать как артефакты. Ошибочное заключение на этот счет указанных авторов связано с тем, что они судили о направлении движения ресничных токов по направлению движения тяжелых частиц взвеси корунда. Однако, осев на поверхность мукуса, эти частицы могут двигаться вместе с ним в сторону рта даже в том случае, если ресничные токи вода идут в противоположном направлении. Это и вызывало видимый эффект движения захваченных слизью частиц в сторону рта, который был описан как эффект обращения направления биения ресничек в присутствии пищи.
И все же в главном Янг был прав: ресничный аппарат действительно выполняет важную функцию в захвате пищи полипами большинства кораллов, хотя механизм этого участия оказался иной. Он состоит на самом деле в том, что полипы таких кораллов, будучи даже в малоактивном состоянии, а иногда и будучи вообще закрытыми, выпускают изо рта слизевые тяжи. Эти тяжи вытягиваются между щупальцами вдоль септ. Под действием достаточно сильных и постоянных ресничных токов вода они "взбиваются", образуя распушенную сеть клейких нитей. Течения, образуемые ресничными токами, взаимодействуя со слизевыми сетями, образуют эффективный фильтрационно-седиментационнгй аппарат (Jorgensen, 1966). Этот механизм полипы кораллов используют не только для фильтрации питательной взвеси (бактерий, псевдопланктона) их водной толщи, но и для хищного питания, а также для ловли малоподвижных жертв, таких, как представители слизистого планктона, адиендикулярии, разнообразные личинки, яйца, цисты. Оба эти механизма ловли и заглатывания животной пищи с помощью книдома и щупалец и с помощью слизи работают одновременно и взаимосвязанно. Они активизируются одним и тем же пусковым триггером, имеющим химическую природу, который приводит полипы в состояние пищевой реакции.
Состояние пищевой реакции наступает, когда добыча находится вблизи полипа или входит с ним в прямой контакт. Это явление было описано уже первыми исследователями пищевого поведения актиний и кораллов (Carpenter, 1910; Vaughan, 1912). Они отмечали, что при добавлении в аквариум с кораллом зоопланктона кусочков мяса или даже вода, в которой ранее сидел зоопланктон, полипы немедленно проявляют пищевую реакцию. Они вылезают из своих кораллитов, их щупальца вытягиваются и приходят в интенсивное движение, околоротовый конус вытягивается, образуя "губу". Полип начинает периодически раскрывать рот, втягивая внутрь выходящие из него и отходящие от овального конуса тяжи слизи, и заглатывает их (Yonge, 1930; Abe, 1938; Lenhof, 1968; Reimer, 1972; Porter, 1974; Lewis, Price, 1975). Специальными исследованиями (Mariscal, 1971) показано, что триггером пищевой реакции полипа является воздействие на его хеморецепторы молекул некоторый аминокислот, прежде всего пролина и глютатиона, которые вызывают эту реакцию при очень малых концентрациях: 10-5—10-7 М. Таким образом, для запуска пищевой реакции достаточно присутствия в воде рачков, которые выделяют во внешнюю среду аминокислоты (Johannes et al., 1969). Их экскреция усиливается, когда покровы жертвы пробиваются нематоцистами и в среду начинает выходить содержимое клеток, интенсифицируя пищевую реакцию полипа.
Характерная особенность склерактиниевых кораллов как хищников состоит в том, что размеры их жертв фактически не ограничиваются размером самого полипа. Для этого у полипа имеется ряд приспособлений. У кораллов с малой полостью кораллита и с колонной в ее середине тело полипа в активном состоянии полностью выходит из кораллита. Рот полипа, так же как и его кишечная полость, может в этом случае растягиваться в несколько раз, позволяя ему заглатывать крупные жертвы (Yonge, 1930). У тех кораллов, полипы которых в активном состоянии не поднимаются из полости кораллита, эта полость сама обычно бывает просторной и в ней редуцирована центральная колонна (Euphyllia, Asteropora). У других видов таких кораллов сильно развиты мезентериальные нити, с помощью которых коралл может переваривать крупные жертвы снаружи, не заглатывая их в кишечную полость. К последним можно отнести такие массовые склерактинии, как Pashyserias, Montipora, Galaxea, Stylophora, и некоторые меандрины (Merulina). В соответствии с наблюдениями Янга коралл Pocillopora с мелкими полипами размером 1-1,5 мм может заглатывать зоеа втрое крупнее своей собственной величины. Полипы этого коралла заглатывали даже небольших саггит длиною до 10 мм всего за 30 с, значительно увеличиваясь в размерах. У коралла Leptastrea кораллиты имеют диаметр около 2 мм. Полипы, которые выходят из этих кораллитов, в активном состоянии имеют диаметр 4-5 мм и способны заглатывать большие куски мяса.
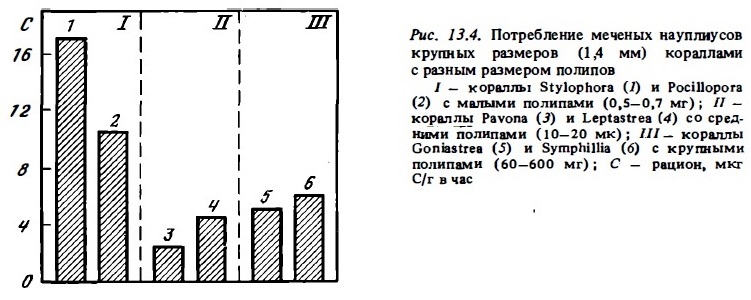
С тем, чтобы выяснить влияние величины жертвы на эффективность хищного питания кораллов с разными полипами, мы выполнили опыты, в которых кораллам с неодинаковыми размерами полипов предлагались крупные науплиусы артемий — около 1,4 мм (рис. 13.4). Оказалось, что кораллы с малыми полипами (около 1 мм) ловят и переваривают крупные науплиусы даже более интенсивно, чем кораллы с крупными полипами.
По механизму питания кораллы делятся на несколько групп в зависимости от преобладания у них одного из упоминавшихся выше механизмов поимки жертв при хищном питании: с помощью щупалец, вооруженных нематоцистами, и с помощью ресничного аппарата и слизевых сетей (Yonge, 1930; 1940; Schmidt, 1972; Lewis, Price, 1975). К первой группе можно отнести некоторые кораллы с мелкими полипами, такие, как Pocillopora, Stylophora, Seriatopora, Madracis. У этих кораллов слизевые тяжи как таковые вообще не вырабатываются и в добыче жертв не используются. Слизь секретируется ими в виде хлопьев. Полипы этих кораллов ловят зоопланктон с помощью щупалец, вооруженных нематоцистами. Пойманные жертвы захватываются венчиком щупалец в "кулачок" и заглатываются полипом. Мелкие планктонные животные, убитые книдоцилем ценосарка дли участков тела полипа, расположенных вне сферы действия щупалец, переносятся в сторону рта щупалец восходящими по колонне полипа вверх ресничными токами (Yonge, 1930).
Другая группа кораллов, имеющая мелкие полипы с очень короткими щупальцами и плоские ребристые колонии (Agaricidae), и ловит добычу вообще практически без участия щупалец с помощью слизевых сетей, обильно выбрасываемых полипами изо рта и сходящими с их оральных конусов. У некоторых видов этой группы в венчике коротких щупалец на каждом полипе имеется 1—2 удлиненных щупальца-дворника, которые совершают интенсивные движения, когда полип находится в активном состоянии, и ловят добычу. При стимуляции полипа такого коралла добавкой пищи он проявляет ярко выраженную пищевую реакцию: широко разевает рот, втягивает в него выпущенную им слизевую сеть, после чего рот закрывается и полип некоторое время переваривает добычу (Lewis, Price, 1975). У этой группы кораллов важное значение как в поимке, так и в переваривании добычи вне их кишечной полости играют мезентериальные нити, особенно сильно развитые у коралла Pashyseris, лишенного щупалец (Yonge, 1930).
Наиболее многочисленная среди склерактиний третья группа кораллов, полипы который используют оба описанных способа хищного питания. Она представлена преимущественно кораллами со средними и крупными полипами, имеющими слабоветвящиеся, массивные или плоские колонии. К этой группе относятся: Acropora, Favia, Syderastrea, Mussa, Isophyllia, Meandrina, Dendropyra, Gala.xea, Pavona, Hydrophora и другие родственные им родит Почти все они имеют полипы с широким оральным диском и длинными щупальцами. В активном состоянии они полностью выходят из кораллита и широко распукают свои щупальца, так что часто под густой их сетью не видно самой колонии коралла. Рот полипов при этом приоткрыт, его края отвернуты, как бы образуя губу. От орального диска в сторону септ идут сильные ресничные токи. Изо рта полипа выходят тяжи слизи, разбиваемые ближе к краям септ на бахромчатые слизистые сети интенсивными ресничными токами. В присутствии пищевого раздражителя, скажем при добавке рыбного сока или науплиусов артемий, наступает резкая пищевая реакция. Рот широко раскрывается, слизь втягивается в рот и заглатывается. Щупальца начинают активно двигаться, пытаясь схватить жертву. Если добыча поймана щупальцем, она немедленно отправляется им в открытый рот. Иногда ротовой конус вытягивается и рот сам "снимает" добычу со щупальца. Щупальце может также положить добычу на слизевый тяж, который затем втягивается в рот и заглатывается вместе с добычей. Таким путем заглатывания добычи часто пользуются акропоры и Hydnophora. У многих кораллов этой группы, например у Montastrea, также имеют удлиненные щупальца-дворники, которые находятся в интенсивном движении и весьма эффективно ловят добычу.
Одно из свойств склерактиниевых кораллов как активных хищников состоит в их способности быстро и эффективно переваривать любую проглоченную или просто захваченную ими добычу. Уже первыми исследованиями об их питании было установлено, что основными органами пищеварения у кораллов являются мезентериальные нити (Matthai, 1973; Boshma, 1925; Yonge, 1930а, b; Yonge, NichUos, 1931). Это довольно сложно устроенные органы мезентерия, которые на своих концах имеют ресничный эпителий, книдоциль и железистые клетки. У основания мезентериальной нити имеется зона абсорбции, которая в большом числе содержит фагоцитирующие клетки. Эта зона является также и основной зоной мезентерия, где происходит экскреция неусвоенных продуктов пищеварения (Yonge, 1935, 1940; Goreau, Philpot, 1956). Процесс пищеварения у кораллов включает два этапа: воздействие экзоферментов-протеаз и столь характерный для кишечнополостных фагоцитоз. Попадание проглоченной жертвы в кишечник немедленно вызывает снижение рН среды от 7,8 до 7,1. Из секторной зоны мезентерия выделяются мощные протеазы, которые в короткое время размягчают ткани жертвы и гидролизуют белки до полипептидов. Одновременно мезентериальные нити совершают активные движения в целентероне и дробят размягченные протеазами-пептидазами ткани жертвы на мелкие частички. Эти частички тотчас же захватывают фагоцитирующие клетки и переносят их в ткани полипа. Блуждающие клетки разносят пищевые частицы клеткам тканей полипов, в которых они окончательно перевариваются внутриклеточно. Неусвоенные остатки пищи транспортируются в зону экскреции, находящуюся на мезентериальных нитях, и там выделяются обратно в кишечную полость, из которой они выбрасываются наружу и сбрасываются ресничными токами за пределы полипа (Yonge, Nichals, 1931; Yonge, 1940). Набор протеаз и их характеристики могут варьировать у разных групп кораллов. Установлено также, что у некоторых корадлов имеются ферменты, которые гидролизуют животный гликоген.
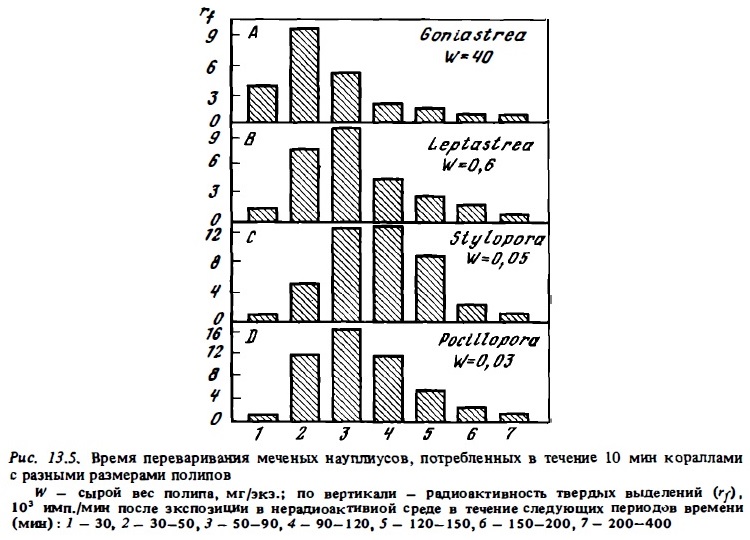
Длительность переваривания животной пищи кораллами зависит от величины проглоченной жертвы. В среднем в соответствии с результатами визуальных наблюдений время между заглатыванием рачка и выбросом его пустой шкурки полипом составляет 1,5-4 ч (Boschma, 1925; Yonge, 1930b; Yonge, Nicholls, 1931; Porter, 1974; Coffroth, 1984). Согласно данным Портера (Porter, 1.974), продолжительность переваривания науплиусов артемий размером 1—2 мм кораллом Montastrea составляет 1,5-2 ч. Переваривание кусочков мяса и крупных зоопланктеров, соизмеримых с размерами своих полипов, может затягиваться до 10—12 ч. Мы попытались определить время переваривания разными кораллами с помощью радиоуглеродного метода. Результаты опытов, представленные на рис. 13.5, показывают, что для кораллов с крупными полипами время переваривания близко к 2—2,5 ч и у кораллов с мелкими полипами — 2,5-3 ч. Усвояемость рачков полипами определилась с помощью радиоуглеродного метода. В среднем она оказалась близкой к 70-80% (Сорокин, 1984в).
Склерактиниевые кораллы способны к фильтрационно-седиментационному питанию. Установлено, что они могут отфильтровывать из воды планктонных бактерий (Di Salvo, 1971а, 1973; Сорокин, 1971, 1972; Sorokin, 1973, 1978а), псевдопланктон, детрит (Lewis, Price, 1975, 1976) и нейтральную взвесь (графит, Lewis, 1977а). Механизм фильтрационного питания кораллов был подробно изучен последними авторами. Он сходен с аналогичным механизмом у многих фильтрующих беспозвоночных (Jorgensen, 1966) и состоит в следующем. С помощью ресничного эпителия животное создает токи воды, проходящие над слизевыми тяжами или сетями. Пищевые частицы, содержащиеся в воде, - бактерии, водоросли, детрит, мелкий зоопланктон, фекалии — захватываются слизью. В этом захвате частиц, особенно коллоидных, не последнюю роль, по-видимому, играют электростатические силы, поскольку слизь, будучи муко-полисахаридом, содержит положительно заряженные центры в виде аминогрупп, в то время как мицеллы органических коллоидов и частицы взвеси могут нести заряд противоположного знака, имея на своей поверхности радикалы карбонатных кислот.
Полипы кораллов используют имеющийся у большинства из них фильтрационный механизм практически беспрерывно в течение суток, независимо от суточной ритмики их активности. Находятся ли полипы в активном состоянии или в неактивном, когда они зарыты и спрятаны в кораллиты, они выпускают изо рта и с поверхности своего орального конуса тяжи слизи. Расползаясь по поверхности полипа, они подхватываются ресничными токами и вытягиваются вдоль септ, взбиваются в бахрому, образуя слизевые пряди и сети (Coles, Strathman, 1973). Взаимодействуя с этими сетями, частицы взвеси, гонимые ресничными токами, вылавливаются ими и постепенно аккумулируются на их поверхности. Если частицы содержат мало питательного материала, они, накапливаясь, обрываются с хлопьями слизи и сбрасываются ресничными токами. Наоборот, если среди отловленной взвеси достаточно питательного материала, он вызывает пищевую реакцию. Полип раскрывает рот, втягивает пишевой комок вместе со слизью и заглатывает его (Lewis, Price, 1975). Такую пищевую реакцию полипа можно вызвать и днем, когда полип находится в неактивном состоянии и прячется в кораллите. При добавке пищевого стимулятора он раскрывает рот, втягивает и заглатывает слизевые тяжи с осевшей на них пищей.
В числе кораллов, которые наиболее интенсивно продуцируют слизевые сети, Montastrea, Mussa, Diploria, Colpophyllia, Lobophyllia, Hydnophora, Pavona, Acropora. В то же время некоторые кораллы, в том числе и те, для которых показана в эксперименте способность питаться взвешенной пищей (бактериопланктоном), вовсе не образуют слизевых тяжей. Они секретируют очень мало слизи в виде отдельных хлопьев, используя ее для обездвижения пойманных жертв и для облегчения их заглатывания, окутывая их слизью (Abe, 1938). К таким кораллам относятся Pocillopora и Porites (Lewis, Price, 1975), кораллы, по-видимому, имеющие несколько отличный механизм седиментационного питания. Ресничные токи у этих кораллов идут не от рта к ценосарку, а, наоборот, от ценосарка вверх по колонне к щупальцам и ко рту, куда они и подносят взвешенный материал. Последний улавливается хлопьями слизи и формирует пищевой комок. Другой способ питания взвешенной пищей (например, бактериями) у таких кораллов состоит в переваривании осевших на их эпителии пищевых частиц протеазами, которые секретируются на всей его поверхности. После предварительного гидролиза белков глобулы образовавшихся из них полипептидов подвергаются фагоцитозу блуждающими клетками, переносятся ими в ткани полипа и там перевариваются внутриклеточно (Wfon, 1975). По мнению ряда исследователей, клетки эпителия полипов способны захватывать бактерии, оседающие на их поверхность, путем пиноцитоза, поскольку по всей поверхности эктодермы полипа размешены мембранные инвагинации, которые функционально приспособлены для пиноцитоза — захвата клетками частиц путем втягивания мембраны в цитоплазму (Goreau, Phippot, 195 6; Goreau et al., 1971).
Способность кораллов потреблять из вод находящиеся в ней я очень малых концентрациях меченые С1 4 сахара была показана ранее Стефенсом (Stephens, 1962). Этот автор сделал вывод о том, что кораллы могут потреблять из морской вода растворенные в ней низкомолекулярные органические соединения. В связи с этим Томас и Нора Горо (Goreau, Goreau, 1960а; Goreau et al., 1971) предприняли исследование судьбы поглощенного полипом меченного тритием лейцина. Оказалось, что уже спустя час после окончания процедуры мечения радиоактивный лейцин обнаруживался на радиоавтографах срезов во всех клетках ткани его эпителия. Спустя сутки метка распределяется равномерно во всех тканях полипа, что доказывает истинное усвоение полипом потребленного из морской воды растворенного органического вещества. Эктодерма полипов имеет необычайно сложное строение. Она покрыта микроворсинками, мембранными образованиями, инвагинациями плазмалеммы (Coreau, Philpot, 1956). Это в огромной степени увеличивает контакт поверхности эпителиальных клеток с внешней средой, облегчая потребление молекул РОВ из вода. При этом потребление крупных коллоидных частиц пищи происходит путем пиноцитоза, а потребление мелких молекул РОВ осуществляется путем активного ферментного транспорта (Goreau et al., 1971). Это было доказано нами экспериментально в опытах с кораллом Pocillopora. Опыты показали, что потребление индивидуальных РОВ весьма специфично, что указывает на участие ферментов-переносчиков в этом процессе (Сорокин, 1977).