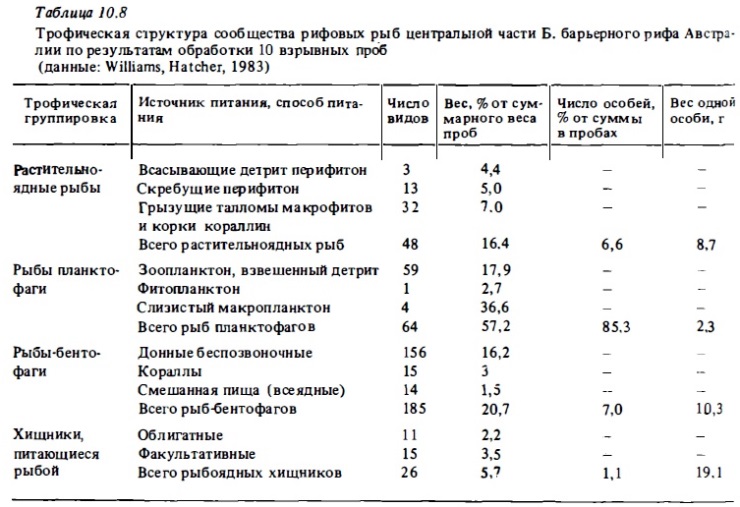
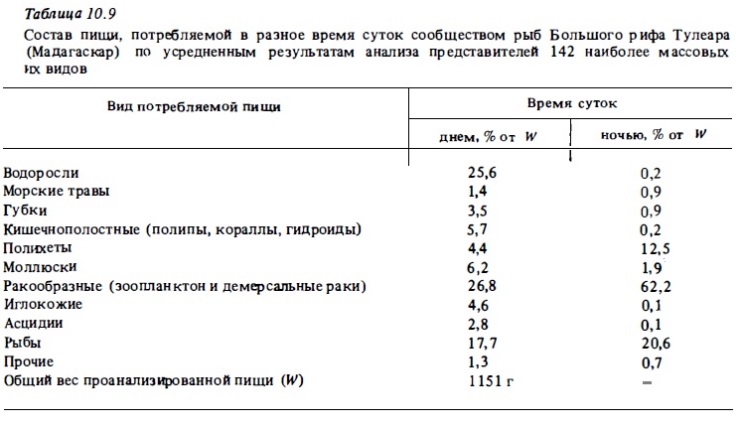
Анализ данных о питании рифовых рыб и соотношении трофических группировок дал возможность выявить некоторые принципиальные отличия ихтиофауны рифа от ихтиофауны прибрежных биотопов умеренных вод. Они состоят главным образом в том, что в сообществах рифовых рыб большой процент составляют растительноядные ах виды (см. табл.10.7). Как по числу видов, так и по численности и биомассе (см. табл.10.8) растительноядные рыбы находятся в числе наиболее массовых трофических группировок (гильдий) рыб. В прибрежных биотопах умеренных вод, где общая ихтиомасса рыб и общее число их видов в среднем в 5-10 раз ниже, чем на рифе, истинно растительноядные рыбы как таковые практически отсутствуют. Доля растений в рационе рыб умеренных вод измеряется считанными процентами (Bakus, 1967), тогда как в зоне коралловых рифов она может измеряться десятками процентов (табл.10.9). Вполне очевидно, что столь значительное участие растительноядных рыб в использовании автотрофной продукции укорачивает пищевую цепь. При этом существенно (примерно вдвое) возрастает эффективность использования основных энергетических ресурсов экосистемы рифа.
Другая характерная особенность трофической структуры сообществ рифовых рыб, отличающая ее от сообществ рыб прибрежных биотопов умеренных вод, состоит в обилии в их составе рыб-планктофагов, питающихся зоопланктоном. Это преимущественно мелкие донные рыбы, живущие в убежищах рифа, а также пелагические личинки рыб и мелкие пелагические планктофаги. Их доля в общем числе видов и в общей биомассе рыб чаще всего соизмерима и даже иногда превышает долю рыб-бентофагов (см. табл.10.8). Так, во взрывных пробах рыб, собранных в центральной части Б. барьерного рифа, более половины особей по весу и около 85% по численности составляли мелкие рыбы-планктофаги со средним весом 2,3 г. Рыбы-планктофаги, выедая зоопланктон из проходящих над рифом океанских вод, обеспечивают поставку внешней энергии и биогенов в экосистему рифа. Эта дополнительная энергия поддерживает существование многих других видов рифовых рыб. Как видно из табл.10.8, число видов рыб-планктофагов достигало четверти всех видов рыб на рифах Б. барьерного рифа. На атолле Эниветок на их долю приходилось 15% всех видов и 32% общей численности рыб.
С учетом же личинок и молоди доля рыб-планктофагов может быть значительно выше.
Внутри семейств и даже родов типично рифовых рыб с большим числом видов имеется тонкая специализация в отношении способов питания. и вида потребляемой пищи (Bakus, 1957). Эта специализация закреплена в виде морфологических и поведенческих адаптаций. В числе семейств, отличающихся наиболее разнообразной специализацией в отношении использования ресурсов рифа: Pomacentridae, Chaetodontidae Gobiidae. Так, в составе помацентрид есть растительноядные виды, питающиеся макрофитами и перифитоном. Есть среди них планктофаги, бентофаги, всеядные виды, рыбы-чистильщики. Каждая из этих трофических группировок помацентрид имеет соответствующую морфологию ротового и двигательного аппарата, раскраску тела и соответствующих тип поведения (Emery, 1973). В составе рыб-бабочек из рода Chaetodon часть видов питается полипами только склерактиниевых кораллов. Другие виды этого рода предпочитают полипы альционарий. В составе этого рода есть также бентофаги и всеядные виды, одни из которых питаются преимущественно бентосом, а другие используют растительную пищу — макрофиты и перифитон. При этом трофической их специализации соответствует свой тип строения ротового аппарата (Harmelin-Vivien, 1981; Reese, 1981).
Примером высокой специализации близких видов рифовых рыб являются: Acanthemblemaria spinosa, А. greenfeldi. Обе рыбки очень похожи внешне и живут в норках. Оказалось, что оба вида различаются по спектру питания (Greenfield, Greenfield, 1932). Специализация в потреблении различных макрофитов, растущих в пределах одного биотопа, двумя близкими видами растительноядных сиганид - Aplysia juliana и А. dacty-lomela помогает их совместному выживанию на одной и той же территории (Carefoot, 1970).
Одну из главных трофических гильдий в сообществах рифовых рыб формируют растительноядные их виды (см. табл.10.7). Нередки случаи, когда они по биомассе даже занимают доминирующее значение. В среднем же их биомасса близка к 15-25% от общей ихтиомассы и число их видов составляет 5-20% от общего числа видов в сообществах (см. табл.8). Растительная пища (перифитон, макрофиты) составляет также значительную долю рациона многочисленной на рифах группы всеядных рыб из сем. Acanthuridae, Pomacentridae. В числе причин, обусловливающих столь обильное развитие рыб-фитофагов среди типичных рифовых рыб, можно назвать, во-первых, преобладание на рифах таких видов растительной пищи, которые могут легко перевариваться рыбами: фитобентос, перифитон, кораллин, всевозможные макрофиты с мелкими талломами (Bakus, 1969). Другая причина, возможно, состоит в том, что, обитая в теплых водах, рыбы имеют высокую температуру тела (25-30°С), что ускоряет работу пищеварительных ферментов, перерабатывающих растительную массу. Эволюционно растительноядные рыбы рифа являются одной из поздних ветвей развития костистых рыб рифа. Они произошли от хищных рыб-потомков древних рыб-акантоптеригий в позднем эоцене (Hobson, 1974). Для них характерен маленький рот с мощными челюстями и массивными зубами, которые, часто срастаясь, образуют пластины типа клюва (сем. Scaridae). В числе других их адаптаций можно отметить высокую эластичность желудка, вмещающего большой объем растительной массы, длинный кишечник, наличие глоточных зубов для измельчения грубой растительной пищи и корок известковых водорослей. Среди пищеварительных ферментов у большинства растительноядных рыб (исключая кефалей) отсутствует фермент целлюлоза, разрушающий стенку клеток растений. Возможно, этот процесс происходит у них с помощью ферментов населяющей их кишечник микрофлоры.
По способам питания растительноядных рифовьх рыб делят обычно на ощипывающих ("browsers"), грызущих ("grazers") и сосущих ("suckers") Первые питаются, обкусывая таллом! макрофитов и листья морских трав. Вторые соскребают своими мощными зубами или клювовидными челюстями перифитонные обрастания и корки кораллин. Третьи отсасывают и собирают детритные наилки с поверхности твердых и мягких грунтов, а также собирают верхний слой микрофитобентоса с поверхности донных осадков. Рыбы первой группы имеют тонкостенный желудок, а рыбы второй группы — мощный толстостенный желудок, напоминающий мускульный желудок птиц. Большая длина кишечника и продолжительное время переваривания растительной пищи (до нескольких часов) обеспечивают сравнительно высокую ее усвояемость: от 30 до 60%. Низкая усвояемость некоторых видов растительной пищи компенсируется у растительноядный рыб высокой интенсивностью питания (Ogden, Lobel, 1978).
Таксономический состав растительноядных рыб достаточно широк. В эту трофическую группировку входят целиком 4 семейства рифовых рыб: Acanthuridae, Scaridae, Kyphosidae, Siganidae. Наиболее многочисленны на рифах представители первых двух семейств, которые дают значительную долю общей биомассы рифовых рыб. В состав растительноядных рыб входят также многие виды семейств Pomacentridae, Go-biidae, Chaetodontidae, Blennidae, Balistidae, а также ласкири Diplodus caudomacula. Кефали и рыбы-хирурги Ctenochaetus striatus питаются донными микроводорослями и детритными осадками, значительную долю которых составляют бентосные диатомеи и синезеленые водоросли. Наконец, некоторые полурылы питаются фитопланктоном, собирая его иэ поверхностной пиенки (Hemiramphus dussumeri, Н. brasiliensis). В числе всеядных рыб, в рационе которых значительную долю составляет растительная пища, можно упомянуть также рыб-ангелов Pomacanthus arquatus, Р. paru, кузовков Canthigaster rostrata и спинорогов Monacanthus ciliatus (Randall, 1967).
Растительноядные рыбы проявляют строгую избирательность в отношении потребления разных видов растительной пищи (Hobson, 1974; Ogden, Lobel, 1978; Lobel, Ogden, 1981). Последние авторы в опытах с рыбами-попугаями Sparisoma radians установили, что по степени предпочтения поедаемые ими растения располагаются в таком порядке:
1) трава Thalassia, обросшая перифитоном,
2) трава Halodule,
3) Thalassia без перифитона,
4) трава Syringodium,
5) водоросль Dictyota,
6) водоросль Enteromorpha,
7) водоросль Caulerpa,
8) Halimeda,
9) водоросль Penicillus.
При этом порядок предпочтения определяется прежде всего питательностью данного растения и его токсичностью для данного вида рыб. В числе водных растений, предпочтительно выедаемых рыбами, морская трава Thalassia, водоросли Enteromorpha, Ulva, Codium (Randall, 196l; Earle, 1972; Lobel, Ogden, 1981).
Результатом пресса выедания водных растений в эволюционном плане была выработка растениями защитных приспособлений. В их числе — способность к росту в прибойной зоне рифа, где выедание растений рыбами затруднено (кораллины, нитчатки, морские травы). Многие известковые водоросли, армируя талломы известью, затрудняют тем самым выедание их рыбами (Turbinaria, Penicillus, Halimeda). Некоторые водоросли обеспечивают себе защиту от выедания рыбами, вырабатывая токсины или вещества, мешающие пищеварению (например, таннины). Так, из 48 видов водорослей, населяющих шельф островов на юге Японии, 38 имеют токсины, которые убивают мышей и вызывают гемолиз (Hashimoto, Fusetani, 1972). В саргассах накапливается таннин, в водоросли Asparagopsis — кетоны, в каулерпе — токсин каулерпин, в лауренсии — хлориды, в водоросли Dictyopteris — токсин диктиопсрин. Известна, также токсичность многих синезеленых водорослей.
Растительноядные рыбы оказывают большое влияние на таксономическую структуру, расселение и продуктивность донной альгофлоры, что в конечном итоге влияет и на расселение кораллов и донной фауны (Randall, 1961; Bakus, 1967; Ogden, Lobel, 1978; Montgomery et al., 1980; Hatcher, Rimmer, 1985). Согласно наблюдениям Бакуса (1967), вся скальная поверхность флета атолла Эниветок носит на себе следы рыб-попугаев, которые соскребают с его поверхности перифитонные обрастания, корки кораллин и сгрызают талломы макрофитов. Там, где выедание водорослей затруднено, как правило, наблюдается пышный рост макрофитов и кораллин. Регулирующее воздействие растительноядных рыб на развитие макрофитов и морских трав было установлено экспериментально путем установки на дно сеток, ограждающих участки дна рифа от посещения его рыбами (Earle, 1972; Wanders, 1977; Nelson, Tsutui, 1981). Уже через несколько недель можно бьшо заметить значительные отличия в росте донной альгофлоры. На защищенных участках он был намного интенсивнее, чем на открытых (Vine, 1974). Любопытно, что внутри клеток угнетался рост кораллов и кораллин, поскольку при исключении выедания водорослей рыбами их поверхность быстро покрывалась растительностью, прежде всего нитчатками (Wanders, 1977; Vine, 1974). Согласно наблюдениям многих авторов, выедание водной растительности рыбами является одним из главных условий успешного поселения и роста кораллов и в конечном итоге — одним из главных условий стабильности структуры и роста современных коралловых рифов (Birkeland, 1977; Potts, 1977; Brock, 1979; Reese, 1981).
Пресс выедания донной растительности рифа рыбами заметно снижается на участках рифа, являющихся территорией некоторых рыбепомацентрид, которую они охраняют от других рыб, а также от крупньк растительноядных беспозвоночных (Vine, 1974; Potts, 1977; Kaufman, 1977). В их числе помацентриды: Eupomacentrus platifrons, Е. lividus, Е. nigrificans. Эти небольшие рыбы отважно прогоняют со своей территории не только конкурирующих растительноядных рыб, включая крупных рыб-попугаев, но и удаляют ежей-фитофагов (Diadema). Рыбка обкусывает длинные иглы ежа, потом берет его ртом за игла и оттаскивает за пределы своей территории. В зонах, охраняемых помацентридами, наблюдается более плотный покров макрофитов, чем вне этих зон (Williams, 1980). Установ ка клеток на их территориях практически не влияет на рост альгофлоры (Montgomery et аl., 1980). После удаления помацентрид с их территории гтотность водорослей на ней заметно убывает уже спустя несколько дней. В пределах охраняемых ими территорий обильный рост водорослей препятствует поселению кораллов и кораллин (Lobel, 1980).
Важная функция растительноядных рыб, и прежде всего рыб-попугаев и рыб-хирургов, состоит также в продукции рыхлого известкового материала и в биоэрозии рифа.
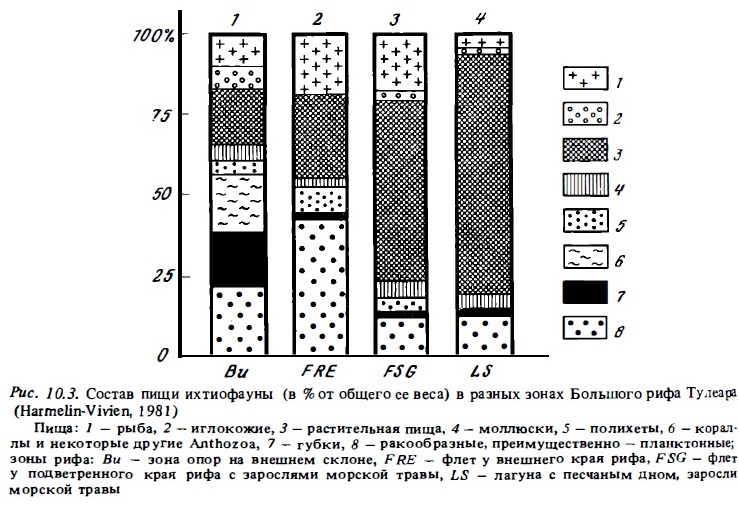
Для сообществ рифовых рыб характерно обилие рыб планктофагов. Судя по числу семейств (более 20) и видов в сообществах (от 5 до 20% от общего числа видов рыб), по общей численности особей (до 85%), по доле в общей биомассе (20-50%, табл. 10.8), а также по доле зоопланктона в суммарном суточном рационе сообщества рифовых рыб (до 43%, рис. 10.3), рыбы-планктофаги представляют собой одну из главных трофических групп в составе ихтиофауны рифа (Randall, 1967; Harmelin-Vivien, 1981; Williams, Hatcher, 1983). Зоопланктон является дополнительным источником питания многих рыб-бентофагов и всеядных рыб (Randall, 1967; Goldman, Talbot, 1976). К тому же почти все рифовые рыбы имеют пелагических личинок, которые питаются зоопланктоном. Рыбы-планктофаги, которые нагуливаются днем, в зоне интенсивного течения используют в качестве пищи преимущественно голоиланктонных копепод, личиночной планктон (меропланктон), икринки рыб. Некоторые виды дневных планктофагов питаются преимущественно сагиттами и слизистым макропланктоном: гребневиками, медузами, сифонофорами, сальпами, которые заносятся теченьем или живут в водах лагуны. В их числе луцианы (роды Caesio, Pterocaesio, Ociurus) спинороги, помацентриды (Pornacentrus fuscus). Некоторые сарганы и каранксы (рода Decapterus, Selar) питаются фитопланктоном и гипонейстонным зоопланктоном.
В составе рыб-планктофагов можно выделить две основные группы, различающиеся по своему происхождению: первичных и вторичных рыб-планктофагов.
Группа первичных планктофагов. В ее состав входят представители семейств типичных пелагических рыб океана, которые приспособились к жизни в зоне коралловых рифов. К ним можно отнести группу сельдевых и сельдеобразных рыб из семейства Clupeidae и Dussurnieridae.
Их приспособление к жизни в зоне коралловых рифов выражается главным образом в уменьшении размеров их тела. Это очень мелкие рыбы, которые, как правило, питаются ночью, а днем, собираясь в стаи, "отдыхают" вблизи рифа или в лагуне.
Группа вторичных планктофагов включает представителей многих семейств типично рифовых рыб, большая часть которых относится к хищным окуневым рыбам или произошла от них (Randall, 1967; Davis, Birdsong, 1973; Hobson, Chess, 1978; Williams, Hatcher, 1983). Вторичные планктофаги имеются практически в составе всех главных семейств донных хищных рыб-бентофагов:
Labridae (Clepticus parrai, Microlabrichthys pascalus),
Lutjanidae (роды Emmelichthyops, Caesio, Pterocaesio, Inermia),
Serranidae (Schulzea beta, Serranus tortugarum, Paranthias furcifer),
Pempheridae (Pempheris schomburgkii, Ociurus crisurus),
Pomadasyidae (род Haemulon, Equetus acuminates),
Pomacentridae (род Chrornis, Pomacentrus coeletus, Amblygliphidodon curacao, Dascycllus ret iculatus),
Gobiidae (роды Ioglossus,Ptereleotris, Cottogobius),
Chaetodontidar (Chaetodon sedentaris),
Apogonidae (Apogon gracilis, A.novaeguineae, А. savayensis, A.binotatus),
Holocentridae (Myripristis pralinus, M.jacobius, M.violaceus),
Cynithidae (Amblycirrithes pinos),
Congridae (Nystacichthys halia, Gorgasia sillneri, Taenioconger halis).
От окуневых рыб произошло и семейство Aterinidae, представители которого являются массовыми видами рыб-планктофагов рифа(рода А ter ina, Allanetta, Pranesus).
Рыбы-планктофаги этой группы в процессе эволюции перейти от хищного питания рыбой или бентосом к цитацию мелкими жертвами - зоопланктоном. Такая смена типа питания вызвала характерные адаптивные перестройки их морфологии и поведения. Морфологические изменения ярче всего выразились в особенностях строения у разных семейств, представителями которых стали планктофаги. Челюсти у рыб-планктофагов по сравнении с их ближайшими родственниками — хищниками вытянулись, рот удлинился и стал трубковидным. При раскрытии такого рта кормовой объект засасывается, как пипеткой. Зубы у рыб-планктофагов на челюстях и на глотке стали мелкими, жаберные тычинки более плотнши, все кости черепа стлали тоньше, увеличился относительна размер глаз, тело стало более веретенообразнш, уменьшились общие размеры тела. У некоторых вторичных планктофагов произошедшая перестройка их морфологии оказалась настолько сильной, что, если бы не некоторые характерные признаки, их вполне можно было бы принять за представителей других семейств. Таковы, например, серра-цида Schulzea beta или луцианы из родов Emmelichthylops и Inermia. Описанные адап-тивн1е перестройки были связаны не только с переходом донных окуневых рыб на более мелкие кормовые объекты, взвешенные в толще воды, но и со связанным с этим их переходом к стайному образу жизни, поскольку в стаях снижается пресс выедания пелагических животных хищниками (Davis, Birdsong, 1973). Именно со стайным образом жизни дневных рыб-плаиктофагов, подверженных атакам хищников, связана их окраска, резко отличная от окраски других видов тех же семейств. В ней преобладают у этих рыб светлые тона с горизонтальными темными полосами, которые нарушают ориентировку хищника при нахождении их в стае. Встречаются также голубоватые и синие тона, маскирующие рыб (род Chromis).
Разные виды рыб-планктофагов выработали разные типы поведения. Часть видов — дневные стайные рыбы, нагуливающиеся днем в толще воды у внешнего края рифа в зоне иитенсивных приливно-отливных течений, с которыми приносится зоопланктон Robson, Chess, 1978). Многие из этих видов (особенно представители рода Chromis и Dascyllus) днем образуют стайки вблизи края рифа или у крупных ветвистых кораллов на внешнем склоне или в проточных каналах на флете. В случае опасности вся стайка немедленно скрывается в расщелинах рифа или в ветвях коралла. Ночью такие стайки обычно рассыпаются и рыбки укрываются в индивидуальных убежищах. У таких рыб окраска тела имеет черты, типичные для пелагических планктофагов. При этом в их стаях, которые часто состоят из видов, относящихся к разным семействам, последние имеют сходную покровительственную или дезоориентирующую окраску поверхности тела (Davis, Birdsong, 1973).
Задача рыб-планктофагов, охотящихся в толще воды в дневное время, состоит в оптимальном питании при минимальной вероятности попасть в пасть хищнику. Разные вины планктофагов решают эту задачу по-разному, что отражается в чертах их морфологии и поведения. Быстро пиавающие рыбки сверетенообразным телом, так же как атери-ны или ранки (Haemulon stiarum), питаются в толще воды, собираясь в большие стаи, что значительно снижает успех хищника в атаке. При нападении эти рыбы отрываются от преследователя, делая мгновенный рывок, который повторяет вся стая. Их реакция столь быстра, что при съемке со вспышкой при выдержке 1/30 с все рыбки в стайке успевают эа это время сделать рывок как реакцию на вспышку света и выходят на снимке размазанными.
Относительно медленно плавающие ярко раскрашенные рыбки Chromis и DascyUus охотятся за зоопланктоном, плавая небольшими стаями, растянутыми по вертикали у края рифа или вблизи больших ветвистых кораллов в зонах интенсивных течений. В случае опасности при приближении хищника стайка скрывается в близлежащем убежище. На поведение стайных рыб-планктофагов значительное влияние оказывает скорость течения и освещенность (Stevenson, 1972). При малой скорости течения рыбы в стае теряют ориентацию вплоть до того, что стая рассыпается. Как только возникает течение, рыбы снова собираются в стаю, особи ориентируются против течения и начинают кормиться.
Некоторые виды дневных рыб-планктофагов пользуются другой тактикой охоты. Они стоят неподвижно в потоке течения вблизи своих убежищ и вылавливают проносимый мимо них планктон. Ктаким рыбам относятся прежде всего змеиные или "садовые" угри из сем. Congridae. Они живут большими колониями в норках, высунувшисьдо половины из воды на расстояние до 20-40 см, и вылавливают проносимый течением планктон (Randall, 1967; Hodson, Chess, 1978). Такой же способ охоты используют многие другие рыбы-одиночки, живущие в норках, вырытых или в мягком грунте или в норках полихет и других сверлящих организмов рифа. В их числе бычки, некоторые рыбы-белки (Holocentridae), а также крылатки (Amblyocyrrites pinos). Эти рыбы лежат неподвижно в засаде на выступе скалы или на ветке коралла и маскируются под цвет субстрата. Периодически они делают молниеносные клюющие движения, вылавливая проплывающих мимо планктеров.
Рыбы бентофаги, которые питаются разнообразнами донными беспозвоночными, обычно образуют на рифах самую крупную по числу входящих в нее видов рыб трофическую группировку (см. табл. 10.8) Число их видов (включая всеядных донных рыб) составляет 40-80% от общего числа видов рыб в сообществах. Их доля в суммарной их биомассе значительно ниже (20-30%), что объясняется разреженностью их видовых популяций при необычайно высоком таксономическом разнообразии и высокой степени специализации в отношении кормовых объектов (Goldman Talbot, 1976; Harmelin-Vivien, 1981; Williams, Hatcher, 1983). В разнос зонах рифа суммарный рацион рыб бенто-фагов может варьировать от 15% в лагуне до более чем 50% в зоне флета и на внешнем склоне рифа. В их составе выделяют три основные группы в зависимости от специализации в отношении предпочитаемой пищи:
а) рыбы, которые питаются сидячими крупными донными животными: губками, ассадиями, сидячими полихетами, гидроидами, зоан-тариями, кораллами;
б) рыбы, питающиеся крупными подвижными беспозвоночными с прочным наружным скелетом: крупными крабами, ежами, офиурами, голотуриями, звездами, крупными гастроподами и двустворчатыми моллюсками, хитонами;
в) рыбы, которые охотятся за мелким подвижнш бентосом мягких грунтов и инфауной скального грунта — преимущественно донными ракообразными (мелкими крабами, остра-кодами, стоматоподами, амфиподами), червями. рыбы-бентофаги первых двух групп в заметном количестве потребляют также растительную пищу, а рыбы третьей группы — мелких донных рыб.
Внутри каждой из перечисленных групп имеется определенная специализация отдельных видов в отношении предпочитаемой пищи, что отражается в особенностях строения их черепа и ротового аппарата (Hiatt, Strasbourg, 1960). Впечатляющим примером такой специализации отдельных видов внутри одного семейства являются даниые о таксономическом составе пищи 7 видов рыб-белок (см. Holoc entridae), обитающих на одном из карибских рифов. Но ночные бентофаги, которые питаются донными ракообразными. Оказалось, что каждый вид потребляет раков разных размерных групп и разных таксонов. Перекрытие ниш по составу потребляемой пищи между этими 7 видами не превышало 0,25. При этом каждвш вид обитает только в своей зоне рифа или в своем биотопе данной зоны рифа (Gladfelter, 1981).
В группе рыб, потребляющих сидячий бентос, можно выделить виды, которые питаются преимущественно губками и асцидиями. В их числе спинороги (Cantherines macrocerus, С .pullus), тетрадоны из рода Arothron (Ahis:Jidus, A.mappa), кузовки (Acanthos-tracton polygonus, A.quadricornis), рыбы-ангелы (роды Holacanthus, Pomacanthus, сем. Chaetodontidae), рыбы-бабочки Chaetodon ephippium. Балянусами питаются крупные губан: Lachnolaimus maximus и рыбы-нетопыри (Ogcocephalus nasutus). Среди рыб, питающихся сидячими кишечнополостными — кораллами и зоантариями, отмечается достаточно четкая специализация в отношении потребляемых ими видов этих колониальных кишечиополостных. Так, спинороги Alutera scripta поедают преимущественно силь-нострекающие и прочные гидрокораллы Millepora. Рыбы-сержанты Abudefduf saxatilis питаются полипами зоантарий. Зоантарии, а также венчики щупалец полихет поедают многие виды рыб-бабочек из сем. Chaetodontidae (Birkeland, Neudecker, 1981). В составе этого семейства некоторые виды потребляют в основном полипы склерактиниевых кораллов. Есть в его составе виды, которые предпочитают полипы альционарий или гидроидов. Они имеют соответствующее строение ротового аппарата, которое позволяет им "высасывать" полипы кораллов из стромы (Reese, 1981). Некоторые рыбы питаются слизью кораллов, соскребая ее с живых колоний (Benson, Muscatine, 1974). В числе последних — помацентриды (Chromis careuleus, Pomacentrus sufflavus), рыбы-бабочки (Chaletodon vagabundus). В числе рыб, питающихся коралллами, рыбы-попугаи (Scarus quacamaia, S.croicensis, S.coelestrinus, Sparisoma aurofrenatum), рыбы-хирурги (Acan-thurus chirurgus), помацентриды (Microspathodon chrysurus). В их кишечниках можно найти обломки твердах кораллов и кусочки альционарий (Randall, 1974). Эти рыбы обкусывают кончики ветвистых кораллов. Рыбы-попугаи своими клювами соскребают строму с полипами и слизью с массивных кораллов вместе с известковым материалом. Мягкие кораллы, в частности горгонарии, поедает наряду с рыбами-бабочками также слинорог Alutera scripta.
В числе рыб, питающихся подвижными беспозвоночными с крупным наружным скелетом или прочной туникой, скат-орляк (Aetobatus narinari), рыбы-ворчуны из сем. Pomadasijidae (Halmulon plumeri, Н. macrostomum), морской карась (Calamus), многие вида губанов (сем. Labridae, рода Bodianus, Halichoeres, Hemipteronotus, Thalassoma), спинороги (Balistes vetula), кузовки (Lactophris trigonus), иглотелы-тетрадоны, диодода. Эти рыбы отличаются мощными челюстями и массивным черепом. Они способны дробить панцири ежей и крабов, звезд, гастропод, хитонов своими челюстями и глоточными зубами.
Подвижный бентос (ракообразных, мелких моллюсков, сипункулид, полихет) потребляют электрические скаты, песчаные акулы, мурены, рыбы-белки (сем. Holocentridae), многие груперы (роды Alphestes, Cephalopholis, Epinephelus, Mycteroperca, Pterometopon), каменные окуни (род Ryphticus), многие губаны (сем. Labridac, рода Bodianus, Halicoeres, Thalassoma), крьтатки, луцианы, рыбы-ворчуны из сем. Pomadasyidae (роды Haemulon, Anisotremus, Rhonciscus, Macolor, Gaterin), горбыли (род Equetus), многие помацентрида (Pomacentrus planifrons, Р. partitus), барабули (сем. Mullidae), камбалы (Botus ocellatus, Achirus proropterus), рыбы-бабочки, скорпены, кузовки (Lactophrys triqueter). Основными компонентами рациона этой группы рыб являются полихеты, креветки, мелкие крабы, которые составляют до 80% объема пищевого комка. Общее качество пищи, которое за сутки потребляется рыбами рифа, может достигать на склоне 2-3 г/м2 беззольного органического вещества, а в лагуне — 0,5 г/м2. Таким образом, донные рыбы выедают значительную долю суточной первичной и вторичной продукции донных биотопов (Hatcher, 1981).
Рыбоядные хищники составляют в среднем 10—20% от общего числа видов в сообществах рифовьх рыб (см. табл. 10.8). Эта оценка весьма приблизительна, поскольку рыбы и особенно их молодь и личинки являются существенной частью рациона многих рыб-бентофагов и планктофагов. Так, в составе ихтиофауны атлантических рифов из общего числа 212 видов желудки 112 видов содержали остатки рыбы. У 48 видов рыба составляла более 50% объема пищи, и только 10 видов питались исключительно рыбой (Randall, 1967). На Б. барьерном рифе из общей биомассы хищных рыб биомасса облигатных хищников, питающихся только рыбой, составляла всего около трети (см. табл. 10.8). Относительная биомасса хищных рыб обьнно бывает сравнительно небольшой (2—10%), причем наибольшее ее значения отмечены в биотопах внешнего склона и флета. Доля хищных рыб в суммарном рационе также варьирует в разных зонах кораллового рифа: от 5% в лагуне до 20% на внешнем склоне рифа (см. рис. 10.3).
В числе видов облигатных хищных рыб наиболее многочисленны груперы, каранги-дат, мурены и барракуда. Среди груперов в числе облигатных хищников чаще всего встречаются представители родов Cephalopholis, Mycroperca, Epinephelus и среди карангид — вида из родов Caranx, Oligoplites, Seriola, Serranus. В число хищных рыб рифа входят также некоторые луциановые (Lutjanus cyanopterus), мелкие тунда (Eutunnus affinis), камбалы (Bothus lunatus), рыбы-клоуны (Antennaris meticeleatus), макрели (род Scomberonorus), рыбы-иглы (сем. Belonidae), рыбы-флейты (род Auiostomus), рыбы-свистульки (род Fistularia), рыбы-ящерицы (род Synodus), рифовые акулы (рода Carcharhinidae, Orectolobidae). Значительный процент рыбы составляют в рационе многих других рыб-губанов, сарганов, рыб-ворчунов, рыб-кардиналов. Наряду с рыбой даже облигатдае хищники потребляют в значительном количестве головоногих: кальмаров, каракатиц, осьминогов, а также крупных ракообразных, в основном креветок. Выедание рифовых рыб хищниками является важным фактором, регулирующим численность и состав их популяций (Sale, 1980; Talbot et al., 1978).