
В соответствии с результатами наблюдений Гохара, подтвержденных позже Горо (Goreau, 1961; Goreau et аl., 1971), сформировалось представление о том, что ксенииды клавулярии и, возможно, многие другие герматипные альнионарии вообще утратили способность к гетеротрофному питанию и обеспечивают себя энергией за счет своих растительных эндосимбионтов. И все же логически принять такое заключение было бы трудно даже в отношении ксений. В самом деле, почему эта группа мягких кораллов оказалась неспособной использовать гетеротрофные источники питания, которые дают кораллам огромное преимущество, обеспечивая их биогенами и энергией взвешенной и растворенной органики? Ответ могли дать только новые исследования. Такие исследования, выполненные в последнее время, снова дали противоречивые результаты. Анализы содержимого кишечников Alcyonium digitatum мягкого коралла умеренных вод подтвердили прежние данные о его способности ловить мелкий малоподвижный зоопланктон, несмотря на редукцию нематонистов на его щупальцах. (Sebens, Koehl, 1984). В них можно было обнаружить личинок асцидий и фораминифер. В кишечниках полипов некоторых горгонарий отмечено присутствие велигеров моллюсков (Bayer, 1956; Grigg, 1972). Согласно Кинзи, (Kinzie, 1970), горгонария Eunicella clavigera ловит планктонных рачков, хотя большинство других изученных им горгонарий с более мелкими полипами ловили только мелкие частицы детрита, но не могли эффективно питаться зоопланктоном. Было отмечено выедание науплиусов артемий горгонарией Leptogorgia (Leversee, 1976). Полипы горгонарии Pseudoplexaura porosa поедали кусочки меченой С1 4 крысиной печени (Murdock, 1978). Согласно наблюдениям Престона (Preston, 1974), суточная активность горгонарий характеризуется двумя максимумами — дневными и вечерними. Дневной ее максимум, как предполагалось ранее (Wainwriget, 1967), связан с их питанием за счет фотосинтеза зооксантелл, а вечерний — с тем, что горгонарии имеют возможность в это время питаться зоопланктоном в период его вечернего максимума. Многие авторы считают, что сама веерообразная форма большинства агерматипных горгонарий и расположение плоскости вееров перпендикулярно направлению доминирующего течения указывают на их приспособленность к ловле мелких планктонных организмов (Theodor, 1963; Grigg, 1972; Leversee, 1972;, 1976; Kinzie, 1973).
И все же многие авторы, изучавшие питание горгонарий в опытных условиях, склонялись к мнению, что они слабые хищники и способны питаться только пассивно плавающими пищевыми частицами. В экспериментах полипы горгонарий хватали и заглатывали такие взвешенные частицы, как шарики сефадекса, пропитанные бульоном, или цисты артемий, но плохо ловили зоопланктоны. Схваченные и даже проглоченные полипами неуплии обычно выпускались ими целыми и невредимыми (Mariscal, Bigger, 1977; Lasker, 1981; Lasker et al., 1983). В то же время имеются данные, указывающие способность альЦионарий отфильтровывать и переваривать фитопланктон (Roushdy, Hansen, 1961). Установлена способность альционарий переварить растительный материал - собственные зооксантеллы, избыток которых полипы выбрасывают в свою кишечную полость. В переваривании зооксантелл у ксении Heteroxenia tuscescens участвуют ее редуцированные мезентериальные нити (Schlichter, 1982) В соответствии с данными того же автора (Schlichter, 1982) ксении способны весьма активно усваивать растворенное органическое вещество из морской воды. Он полагает, что этому способствуют постоянные пульсирующие движения полипов, столь характерные для этих альционарий.
Наряду с РОВ важным источником питания для альционарии, по-видимому, является слизь, выделяемая герматипными кораллами. Судя по результатам опытов, выполненных с массовым видом карибских герматипных горгонарий Pseudoplexaura porosa (Coffroth, 1984), горгонарии интенсивно поедают хлопья слизи, выделяемой герматипными кораллами. Интенсивность потребления слизи составляла около 0,9 мг сухого веса на 1 г сухого веса колонии за 10 мин, причем почти 15-30% его усваивалось полипами. Оценивая эти результаты, следует учесть, что калорийность слизи довольно высока: 4-5 кал/мг беззольного сухого веса, причем вещество слизи обогащено азотом (Rublee et аl., 1980). Часовое потребление слизи горгонариями было близким к 3 мг сухого веса вещества на 1 г органического вещества их колоний или около 100 ккал/г в сутки, что соответствует компенсации 120-290% суточных трат на обмен. Продолжительность переваривания съеденных горгонарий Plexaura хлопьев слизи в опытах этого автора была близка к трем часам (см. рис. 13.6).
В относительно недавнее время Люис предпринял попытку решить спорный вопрос о способности мягких кораллов к гетеротрофному питанию и попытался выяснить механизмы захвата пиши их полипами (Lewis, 1982). Исследования были выполнены с 24 видами альционарий, горгонарий, а также с представителями порядка Stolon ifera (Tubipora) и Telestacea (Telesto). Некоторые из этих видов (Sinularia capillosa, Eunicea tourneforti, Paralenmalia digitoformis, Plexaura tlexnosa, Gorgonia ventalina) оказались неспособными ловить и заглатывать жертвы. Остальные, изученные Люисом октокораллы, включая как герматипные их роды и виды (Lobophytum, Sarcophyton, Sinularia, densa, Capnella, Limnalia, Isis, Juncella, Pseudopterogorgia, Plexaura, Telesto, Tubipora), так и агерматипы (Dendronephthea, Rumphella, Briareum, Muriceopsis, Gorgonia), ловили и заглатывали планктонных рачков. Только ксенииды отвергали в лабораторных опытах все виды взвешенной пищи, как это было ранее показано Гохаром (Gohar, 1940). Однако, анализируя содержимое желудков ксений, взятых из природы, Люис выявил у части из них проглоченных полипами планктонных организмов с размерами до 1,5 мм: рачков, полихет, велигеров, яиц беспозвоночных. Неспособность ксений ловить взвешенную пищу в опытных условиях в стаканчиках, помещенных на столик микроскопа, возможно, объясняется тем, что для успешной их ловли длинными щупальцами со слабой мускулатурой последние должны делать пассивные возвратно-поступательные движения под воздействием движений воды на мелководьях рифа, где они обычно обитают. Именно этим, по-видимому, и объясняется отсутствие ксений в приглубых участках склона и лагуны.
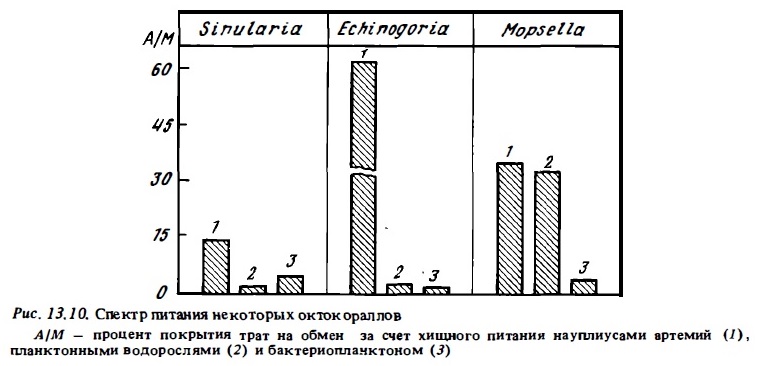
Противоречивость существующих представлений о способности октокораллов к гетеротрофному питанию связана также и с тем, что они гораздо хуже, чем склерактинии, переносят условия эксперимента (Lewis, 1982). К тому же стратегия питания и его механизмы сильно варьируют в пределах этой группы. Мы попытались подойти к решению данного вопроса путем определения спектра питания представителей разных таксонов октокораллов с помощью радиоуглеродного метода, создавая акклимированным к условиям эксперимента колониям весь спектр потенциальных источников питания в одной серии опытов, исключив тем самым влияние на результаты их физиологического состояния. Параллельно в тех же сериях опытов определяли спектр питания склерактиний в целях сравнения их с октокораллами. Типичные результаты опытов, выясняющих спектр питания октокораллов, приводятся на рис. 13.10. В целом они подтвердили вьгаоды Люиса (Lewis, 1982) о способности октокораллов к гетеротрофному питанию. Однако спектры питания у представителей разных их групп оказались весьма различными. У большинства подопытных видов при оптимальной концентрации пищи (2-4 мг/л сырой биомассы) преобладало хищное питание. На втором месте по интенсивности усвоения было питание их бактериопланктоном. В меньшей степени большинством из них потребляются и усваиваются бактериопланктон и РОВ. Наиболее активными хищниками среди изученных видов октокораллов оказались агерматипные октокораллы: альционарии (Dendropephtea, Spongotes) и горгонарии Bebryce, Acabaria. В питании горгонарий с мелкими полипами (Mopsella) значительную роль играли водоросли, которые переваривались и усваивались полипами. Способность мягких кораллов в отличие от склерактиний переваривать растительные клетки была ранее показана Гохаром (Gohar, 1940). Все изученные октокораллы потребляли и усваивали бактериопланктон и растворенное органическое вещество. Установлено, что в процессе потребления РОВ мягкими кораллами (Xenia) участвует ферментная система активного транспорта (Schlichter, 1982).