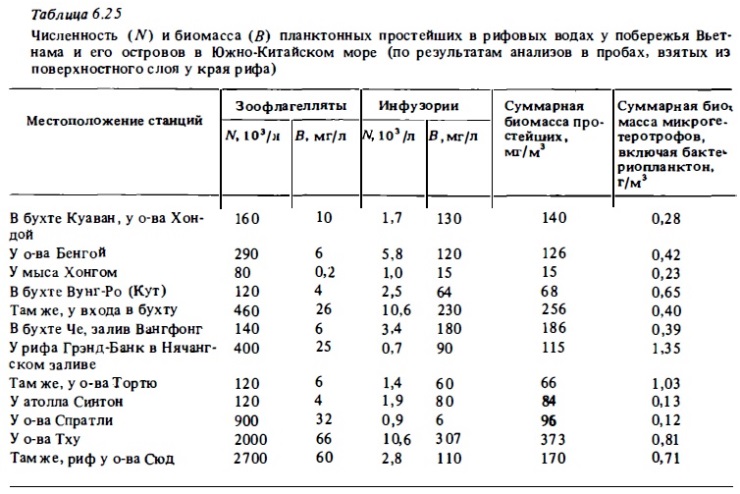
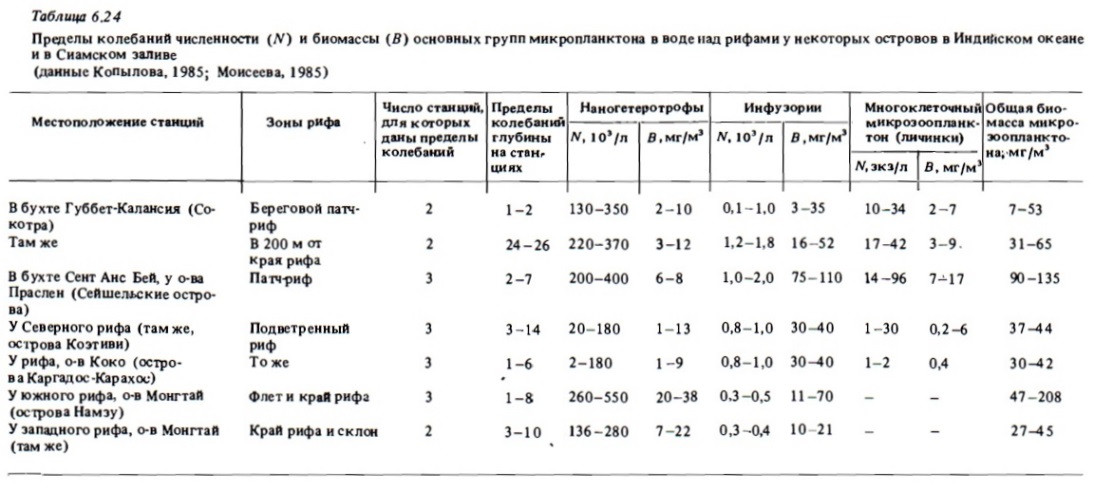
Несмотря на достаточно широкий размах исследований микрозоопланктона открытого океана (Сорокин, 1982), в рифовых водах этот важнейший компонент зоопланктона до недавнего времени практически не изучался, особенно в количественном аспекте. В некоторых работах по планктону рифов есть отдельные данные о содержании и науплиусов, и инфузорий-тинтиннид в сетных пробах зоопланктона (Marshall, 1933); Sammarco, Crenshaw, 1984). Исследования, выполненные в 1981 и 1984 гг. выявили первостепенное значение микрозоопланктона как компонента планктонных сообществ рифовых. вод (Сорокин, Тяпкин, 1984; Копьшов, 1985; Моисеев, 1985). В соответствии с полученными данными доминирующей его группой по биомассе, как правило, являются инфузории (табл. 6.24, 6.25). Численность зоофлагеллят бывает в среднем на 2 порядка выше численности инфузорий. Они входят в состав наногетеротрофов, куда вместе с ними относят планктонных амеб, мелких радиолярий и гетеротрофные бесхлорофильные жгутиковые водоросли. При этом зоофлагелляты обычно составляют около половины общей численности наногетеротрофов в водах коралловых рифов. Последняя бывает достаточно большой и может достигать полумиллиона в 1 л.

Их биомасса, по оценкам А.И. Копьтова (1985), в среднем составляет 2-10мг/м3. Однако в ряде случаев она может быть намного выше - до 20-60 мг/м3. Такие величины были зарегистрированы на рифах у берегов Вьетнама и его островов в периоды нагонов к ним массы синезеленых водорослей.
В составе сообществ зоофлагеллят рифовых вод доминировали рода Monas, Bodo, Oicomonas. В составе инфузорий в водах коралловых рифов преобладали голые их формы родов Strombidium, Strobilidium, Tontonia, Тiarina. Часто в пробах встречались бентопланктонные гипотрихи. Размеры этих инфузорий варьировали в пределах 20-80 мкм. Численность инфузорий в рифовых водах, как правило, была достаточно высокой. В среднем она выражалась величинами 0,3-2 тыс./л (см. табл. 6.25, 6.26).
Биомасса инфузорий в водах рифов в среднем составляла 20-60 мг/м3 (рис. 6.21). Высокая численность и биомасса инфузорий были встречены в воде над патч-рифами в бухте у о-ва Праслен, где наблюдалась вспышка развития фитопланктона. Высокие величины биомассы инфузорий (до 300 мг/м3) были обнаружены в зоне рифов, находящихся в кутовых бухтах у побережья Вьетнама (бухта Че, бухта Ня-Фу), а также у о-вов Тху, где имел место нагон синезеленых водорослей в-зону прибрежных рифов с акватории Южно-Китайского моря (6.26). Величины численности и биомассы инфузорий в водах в зоне коралловых рифов в среднем в 3-10 раз выше тех, которые обычно встречаются в олиготрофных и мезотрафных водах тропической зоны океана. Часто эти величины близки к таковым в наиболее продуктивных водах морских бассейнов.
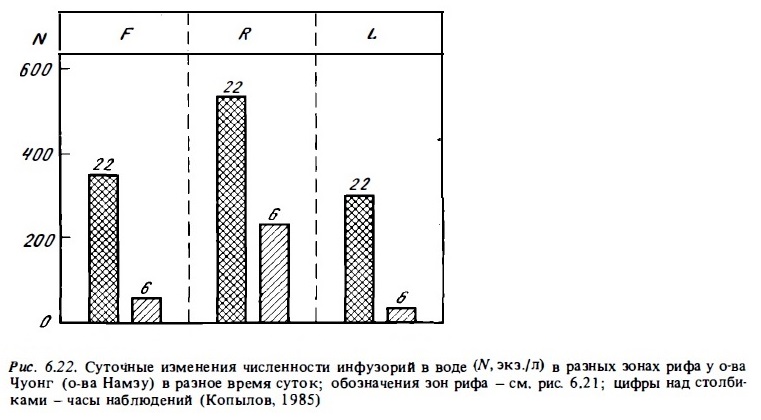
Наблюдения показывают, что, как и численность инфузорий, их биомасса сильно варьирует в разных зонах рифа, в разное время суток и даже в разные дни. Самые низкие ее величины обычно имеют место в мелководных зонах рифа в зоне флета с интенсивным развитием кораллов. Например, в воде над Процветаюищми рифами атолла Синтон биомасса простейших над флетом была в 20 раз ниже, чем у края рифа над глубиной 8 м всего в 20—30м от предыдущей точки (рис. 6.21). Столь же значительными могут быть и суточные изменения численности и биомассы планктонных простейших в одной и той же зоне рифа (рис. 6.22). При этом максимальные величины отмечаются поздним вечерам, а минимальные рано утрам. В числе основных факторов, которые обусловливают столь интенсивную суточную динамику и высокую пространственную изменчивость популяций микразоопланктона в водах рифов, можно назвать выедание, суточные миграции и суточную ритмику размножения. Микразоопланктон, как и фитопланктон, интенсивно выедается донными фильтраторами рифа. Эффект ночного выедания микразоопланктона в зоне рифа можно видеть на рис. 6.22. В течение ночи биомасса инфузорий в воде над рифом убывала в 3-10 раз.
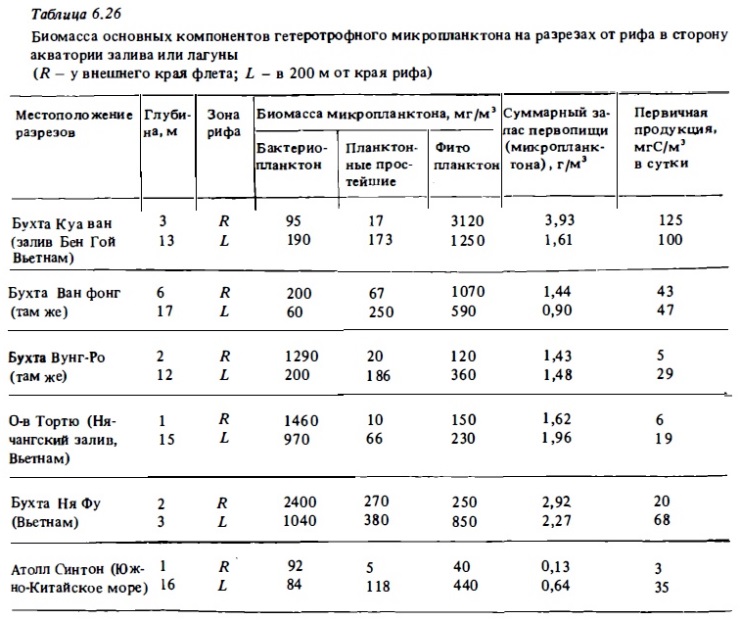
Личиночный или многоклеточный микразоопланктон формирует заметную часть общей биомассы микразоопланктона в среднем от 10 до 20% (см. табл. 6.25). Последняя выражается величинами 20—200 мг/м. Иногда она превышает 300 мг/м3 (см. табл. 6.26). Как видно из этих данных, микразоопланктон является важным компонентом планктонного сообщества. Не подлежит сомнению, что он играет заметную роль
в формировании кормовых ресурсов фильтрующей фауны рифа. Его особое значение следует видеть в том, что организмы микрозоопланктона служат основным источником питания личинок рыб на самых ранних стадиях их развития в тот период их жизни, когда наличие доступной мелкой животной пищи определяет их выживаемость. Важное функциональное значение микрозоопланктона обусловлено тем, что интенсивность его обмена и его удельная продукция в расчете на единицу биомассы примерно в 5-10 раз выше, чем у мезозоопланктона (Сорокин, 1982). Время генерации инфузорий в их естественных популяциях составляет 10-40 ч и время гене рации зоофлагеллят - 7-10 ч (Копылов, 1985). Суточные коэффициенты удельной продукции инфузорий варьировали в пределах 0,4-1,6, зоофлагеллят - 1,5-3,2. Таким образом, в среднем за сутки популяция простейших в рифовых водах воспроизводит свою наличную биомассу. Сравнение на основе этих данных вероятных величин суточной продукции микрозоопланктона с продукцией мезо- и макрозоопланктона в водах рифов показывает, что эти величины соизмеримы. Так, при средней биомассе мезо- и макрозоопланктона в толще вода в суточном цикле 0,5 г/м3 его продукция может оцениваться в 30-50 мг/м3. Средняя продукция популяций микрозоопланктона при ее биомассе 30-50 мг/м3 и при доминировании в ней инфузорий может быть оценена примерно такими же величинами — 30—50 мг/м3 в сутки. Ориентировочные величины дыхания микрозоопланктона в толще вод рифа показывают, что суточное дыхание микрозоопланктона в водах рифов Вьетнама должно было составлять 30—100 мкг О2/л в сутки.