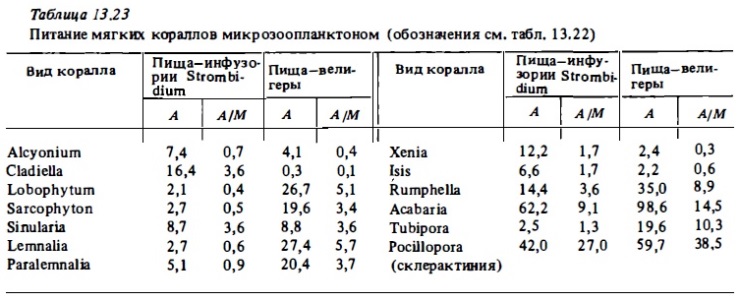
Результаты определений показывают, что все изученные нами массовые представители герматипных октокораллов, как альционарий, так и горгонарий. способны в той или иной мере к гетеротрофному питанию, хотя степень использования ими тех или иных потенциальных источников питания сильно варьирует у разных их представителей. В разной степени отдельные их виды могут использовать практически все основные источники пищи. Они потребляли в опытах мезозоопланктон (науплиусы артемий), микрозооиланктон (инфузорий, велигеров и коловраток; табл. 13.22, 13.23), планктонные водоросли, бактериопланктон и растворенное органическое вещество. За счет хищного питания мезозоопланктоном герматипные альдионарии и горгонарии, судя по результатам опытов, могут компенсировать до трети (10-30%) трат на дыхание. Предположение о том, что они преимущественно питаются микрозоопланктоном не подтвердилось в наших опытах (см. табл. 13.23). Интенсивность питания их коловратками, велигерами и инфузориями в целом была невысокой. За счет их потребления они компенсировали в среднем от 1 до 5% суточных трат на обмен. Среди герматипных октокораллов лучше других потребляли микрозоопланктон Rumphella. Слабее других потребляли его Capnella и Cladiella.
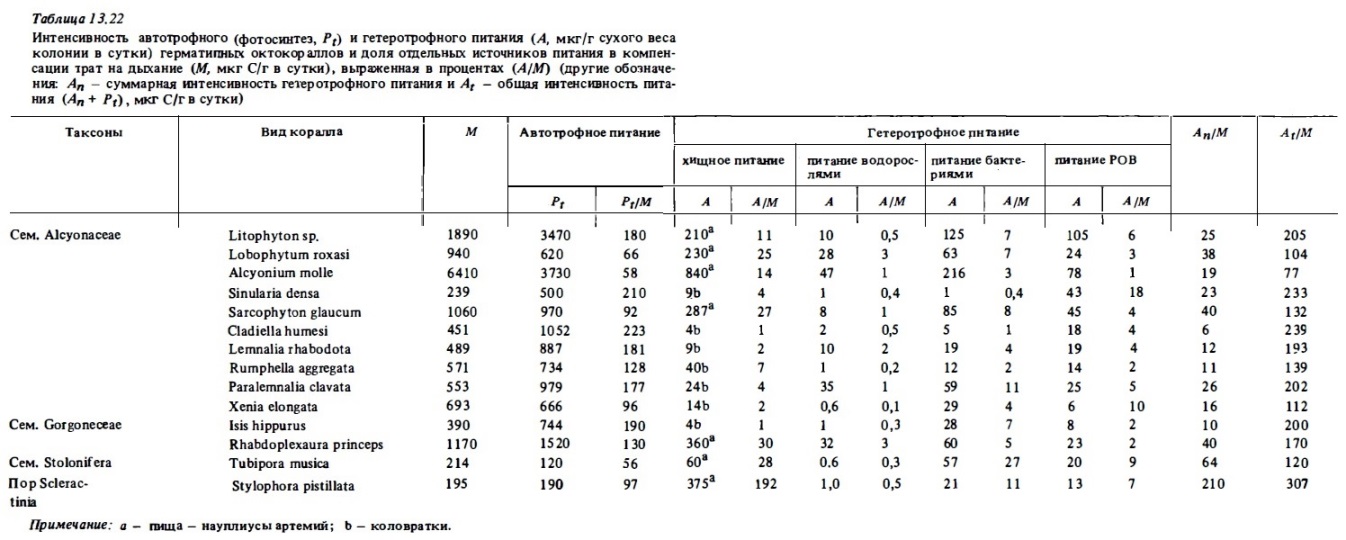
На втором месте в качестве источника гетеротрофного питания у них стоят бактерии. За счет питания бактериопланктоном большинство из них могут компенсировать от 3 до 11% энергетических потребностей. Наибольшей способностью к потреблению бактериопланктона среди октокораллов обладает столонифера Tubipora, компенсирующая за счет его усвоения до трети своих энергетических потребностей. Герматипные октокораллы хорошо потребляют также и растворенную органику, компенсируя при этом 2-14% энергетических потребностей при ее концентрации в воде 0,5-1 мг С/л. Водоросли даже при их достаточно высокой концентрации усваиваются ими в минимальных количествах. Суммарное гетеротрофное питание (см. табл. 13.22) при оптимальной концентрации пищи в воде позволяет разным видам герматипньк октокораллов обеспечивать от 6 до 40% приходной части своего энергетического баланса. Остальную энергию они получают, как это видно из таблицы, за счет фотосинтеза своих зооксантелл. Значение гетеротрофного питания как источника энергии особенно существенно у тех их видов, у которых фотосинтетическая продукция ниже их энергетических потребностей (Pt/Mt < 1). Таковы, например, альционарии Alcyonium, Lobophyton и столонифера Tubipora.
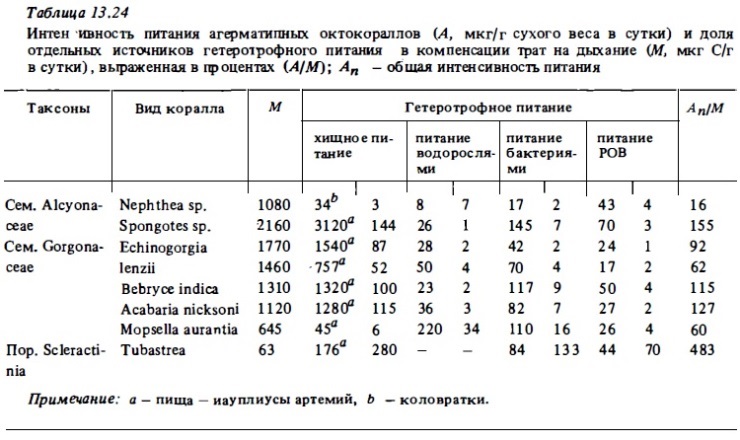
Агерматипные мягкие кораллы оказались более активны, чем герматилные. За счет питания науплиусами артемий или велигерами большинство из них могут компенсировать свои энергетические потребности (табл. 13.24). Самыми активными хищниками оказались альционарии и горгонарии с относительно крупными полипами, такие, как Spongotes Bebryce, Acabarie. В то же время горгонария Mopsella с мелкими полипами практически не потребляла науплиусов, зато с поразительной интенсивностью питались водорослями, а также и бактериями, компенсируя за счет их усвоения до половины трат на обмен. Этим своим свойством она резко выделялась среда других родственным ей и сходным с ней по структуре горгонарий, таких, как Acabaria. Сравнительно слабым хищником оказалась также горгонария Plexauroides. Агерматипные горгонарии способны достаточно интенсивно питаться микрозоопланктоном. Это было установлено опытами с кораллами Acabaria, которая, питаясь велигерами, компенсировала около 15% трат на обмен (см. табл. 13.23). Агерматипная альционария Nephtea питалась микрозоопланктоном (коловратками) намного слабее (3% от трат на обмен, см. табл. 13.24).
Представленные выше данные указывают, на существование тонкой трофической специализации отдельных видов агерматипных горгонарий. По-видимому, некоторые их виды с мелкими полипами ориентированы на потребление разных форм растительного и животного микропланктона, детрита или псевдопланктона. В целом же агерматипные октокораллы в большинстве своем являются хищниками и сравнительно мало используют другие источники взвешенной пищи, а также РОВ, получая за счет использования последних всего 3-6% потребленной им энергии. При сравнении агерматипных видов октокораллов и склерактиний (Tubastrea) в отношении интенсивности и спектра их питания (см. табл. 13.22) выясняется, что последние способы намного полнее и с большей эффективностью использовать доступные источники пищи: зоопланктон, бактериопланктон, РОВ. Так, Tubastrea способна компенсировать энергетические траты, питаясь одним только бактериопланктоном. При оптимальной концентрации основных источников пищи интенсивность усвоения ею пищи почти в 5 раз выше ее энергетических трат.
Представленные выше материалы показывают, что у герматипных октокораллов, гак же как и у герматипных склерактиний, основным источником энергии служит фотосинтез их растительных эндосимбионтов. Вместе с тем эти октокораллы намного слабее, чем склерактинии, пользуются гетеротрофными источниками питания, особенно зоопланктоном. В равных условиях опыта склерактинии намного эффективнее, чем большинство мягких кораллов, использовали зоопланктон, взвешенную пищу и РОВ. Это вполне соответствует данным о более совершенной функциональной морфологии их полипов. Имеющиеся данные позволяют сделать предварительный вывод о том, что высокая конкурентоспособность октокораллов в борьбе за субстрат с гексакораллами, по-видимому, связана не с их трофической активностью и не с оппортунистичностью в отношении источников питания, а скорее с выработанными в процессе их длительной эволюции средствами химического антагонизма (Coll et аl., 1982), а также с их большей толерантностью в отношении к неблагоприятным факторам средам таким, как осушка, загрязнение, затенение (Nishihara, Yamarato, 1974).