
Вопрос о гетеротрофном питании кораллов, о его роли в энергетическом балансе и в бюджете биогенных элементов кораллов давно привлекал внимание исследователей (см. обзоры: Yonge, 1968; 1973; Goreau et al., 1971; Muscatine, 1973). Он вызывал острые дискуссии, в ходе которых высказывались диаметрально противоположные точки зрения: от полного отрицания нужды их в гетеротрофных источниках пищи (Franziskett, 1970) до признания хищного питания кораллов главным и фактически единственным источником энергии, оставляя за зооксантеллами роль вспомогательной системы для удаления продуктов обмена, удержания биогенов и кальцификации (Yопgе, 1940, 1963, 1973). Уже опыты первый исследователей показали, что полипы кораллов активно реагируют на присутствие животной пищи (кусочков мяса или рыбы, рачков), хватают их и отправляют в рот (Duerden, 1906; Vaughan, 1912). В мезентернии пища дробится совместным действием пищеварительных ферментов и механическим воздействием мезентериальных нитей, мелкая взвесь и полипептиды фагонируются и разносятся по тканям полипа блуждающими клетками. Любая растительная пища полипами кораллов отвергается (Jordan, 1907; Vaughan, 1919; Boshma, 1925; Уоngе, Nickholls, 1930а). Однако и тогда было мнение, что, несмотря на способность кораллов к хищному питанию, основную энергию они получают от своих симбиотических водорослей, которыми набиты клетки их эпителия (Gardiner, 1903; Grawier, 1913). Эти вполне резонные взгляды о важной роли зооксантелл как источника питания герматипных кораллов были позже надолго оставлены под влиянием работ известного исследователя физиологии кораллов Янга, выполненных им во время работ Экспедиции Б. барьерного рифа в конце 20-х годов (Younge, 1930; 1930а). Он был решительным сторонником мнения о том, что кораллы являются преимущественно хищниками. Зооксантеллы он считал комменсалами коралловых полипов, основная функция которых состоит в поглощении продуктов обмена полипа, в запасении биогенов, а также в косвенном участии в процессе кальцификации. Его классические исследования питания кораллов не потеряли своего значения до настоящего времени, особенно в отношении способов захвата пищи полипами (Yonge, 1940). Однако его вывод о том, что их энергетика не зависит от фотосинтеза зооксантелл, который он воинственно поддерживал часто против очевидности (до 60-х годов), надолго сдержал развитие представлений о симбиотической природе герматипных кораллов. Появление в60-70-х годах потока публикаций о транслокации фотосинтетической продукции симбиотических водорослей кораллов в ткани полипа хозяина в конце концов окончательно склонило общее мнение исследователей к тому, что все же основой энергетики герматипных кораллов является фотосинтетическая продукция зооксантелл, а гетеротрофное их питание если и имеет для них какое-либо значение, то не как важный источник энергии, а скорее как источник получения биогенных элементов и витаминов из внешней среды. Укреплению представления о подчиненном значении гетеротрофного питания кораллов способствовали также накопленные к тому времени данные о суточном балансе фотосинтеза и дыхания у кораллов. Согласно этим данным, их соотношение (Р/М) у большинства видов склерактиний при достаточной освещенности выше единицы. Это как будто бы позволяло утверждать, что у кораллов автотрофный обмен преобладает и автотрофной продукции одной достаточно для их нормального существования (Johannes, 1974). Но фактически данное утверждение неверно, поскольку траты на обмен включают, Особенно у кораллов, не одно только дыхание. Сюда входят также траты на рост, размножение и, главное, на экскрецию слизи, которая составляет у них 20-40% от суточной продукции их фотосинтеза (Crossland, 1980; Сорокин и др., 1983).
В начале 70-х годов появилась серия работ, показавших, что кораллы способны отфильтровывать, поедать и усваивать планктонный бактерий при их концентрации в воде, близкой к естественной (Di Salvo, 1971а, 1973, 1974; Сорокин, 1971, 1972), в связи с чем Мускатин, подводи итоги исследований по гетеротрофному исследованию кораллов (Muscatine, 1973), высказал мнение о том, что кораллы, по-видимому, наряду с хищным могут пользоваться и фильтрационным способом питания.
Одна из тенденций в изучении питания кораллов, проявившихся в середине 70-х годов, состояла в развитии представлений о том, что огромное разнообразие структур и экологических адаптаций отдельный систематических групп склерактиний отражает их специализацию в использовании доступных им источников энергии в пределах заселяемых ими биотопов рифа. При этом полагали, что такая специализация должна отражаться прежде всего в соотношении автотрофного и гетеротрофного их питания. Такую точку зрения впервые сформулировали Гора и Янг. Они высказались в пользу того, что кораллы с крупными полипами и длинными щупальцами, такие, как Favia, Fungia, Symphillia, являются преимущественно гетеротрофами, питаясь крупными зоопланктоном, будучи сходными в этом отношении с агерматипными кораллами Dendrophyllia. Их зависимость от фотосинтеза зооксантелл, по мнению упомянутых авторов, должны быть минимальной, поскольку они используют ресурсы гетеротрофного питания в большей степени, чем световую энергию. Ресурсы же световой энергии в донных биотопах рифа, занятых кораллами, используют преимущественно кораллы с мелкими полипами, имеющие ветвистые колонии: PociUopora, Stylophora, Seriatopora, Acropora.
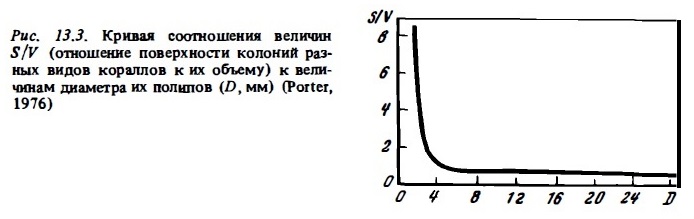
В дальнейшем эта идея была развита в работах Портера (Porter, 1976). Его аргументация в пользу такого деления склерактиниевых кораллов на группу автотрофов и группу гетеротрофов основана на анализе кривой соотношения поверхности колоний к их объему (рис. 13.3). Участок кривой у оси ординат соответствует группе кораллов с мелкими полипами, которые, как видно из кривой, имеют высокую относительную поверхность и соответственно, согласно идеям Горо и Портера, должны быть преимущественно автотрофными. Портер называет этот участок кривой "автотрофным". Участок кривой, соответствующий кораллам с крупными полипами, расположился у оси абсцисс. Он соответствует низкой относительной поверхности колоний, что, по Портеру, характерно кораллов с преобладанием гетеротрофного питания. Это — "гетеротрофный" участок кривой. Внешне описанная теория Портера казалась весьма привлекательной, однако она требовала экспериментального подтверждения.
К концу 70-х годов стало вполне очевидным, что склерактиниевые кораллы способны пользоваться с одинаковой эффективностью механизмами хищного и фильтрационного питания наряду с их способностью использовать в своем метаболизме продукты фотосинтеза зооксантелл (Lewis, Price, 1975; Lewis, 1977; Sorokin, 1978; 1981; Falkovsky et al., 1984). В связи с этим остро встала задача, сформулированная 30 лет назад Одумом и Одумом (1956), состоявшая в определении значения каждого из этих источников питания у кораллов, относящихся к разным систематическим группам и имеющих разную форму колоний и разные размеры полипов. Решение этой задачи помогло бы ответить на вопрос, существует ли в действительности у склерактиний трофическая специализация. Ее решение представлялось также весьма актуальным с точки зрения анализа структуры и функционирования экосистемы рифа. Мы попытались решить эту задачу на примере массовых видов кораллов Б. барьерного рифа Австралии, выполнив исследования их питания и метаболизма на Исследовательской станции университета Квинсленда на атолле Херон в 1975-1976 гг. Результаты этой работы не подтвердили теории Портера о делении кораллов на группы "автотрофов" и "гетеротрофов". Более того, они не подтвердили и их трофической специализации в отношении использования источников гетеротрофного питания. Это говорит в пользу того, что не ресурсы пищи и световой энергии, которых, в биотопах рифа, заселяемых, кораллами, им достаточно, а скорее доступность субстрата, физический стресс и выедание были в числе тех факторов, влияние которых обусловило появление столь разнообразных морфологических форм склерактиний.