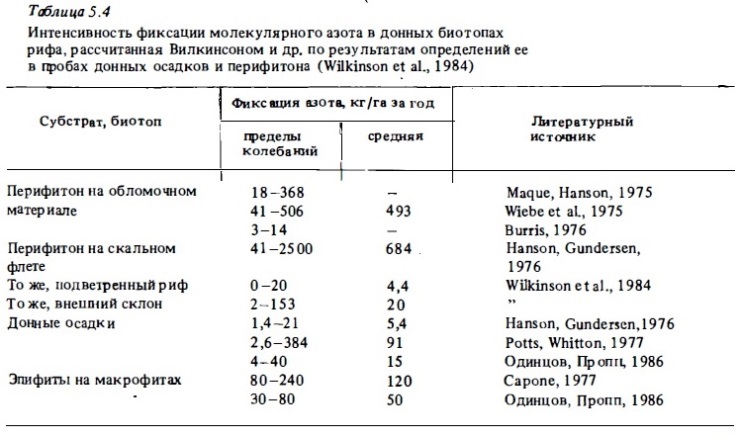
Исходя из найденных в опытах с естественными субстратами величин азотфиксации была вычислена ориентировочная продукция связанного азота из атмосферного в разнотипных донных биотопах рифа (табл.5.4). Ее оценивают величинами 500—900 кг N/га (Webb еt al., 1975). Эти данные, полученные с применением ацетиленового метода, могут быть завышенными в 1,5-2 раза (Burns, 1976). Но даже при средней величине годовой азотфиксации около 100 кг/га в зонах развития перифитона должно производиться около 30 мг N/м2 в сутки. В расчете на водную толщу при глубинах 2—3 м суточная продукция связанного азота выразится величинами 0,2-0,3 мкг ат N/л, что составляет 30-40% от обычного его запаса в рифовых водах.
Долю аэотфиксации в обеспечении донных растительных сообществ азотом можно оценить в 10—30%. Установлено, что процесс азотфиксации усиливается в зонах, где макрофиты выедаются рыбами и их место занимают быстро растущие сообщества перифитона, в которых фиксация азота идет наиболее интенсивно (Wilkinson et аl., 1984). Азотфиксация обнаружена также в телах некоторых губок, имеющих симбиотические синезеленые водоросли (Wilkinson, Fay, 1979).