
В мелководных зонах рифа структуру сообществ в значительной мере контролируют волновой стресс и приливно-отливные течения (Roberts, 1974; Dollar, 1982). Интенсивная гидродинамика сама по себе является необходимым условием нормального существования сидячего бентоса. Она обеспечивает постоянный приток пищи и биогенов, улучшает газовый режим (Hubbard, 1974; Lewis, 1976; Wainwright, Koehl, 1976), а также обеспечивает вынос избытка взвеси, поступающей с берегов в мелководные участки рифа, предотвращая заиление дна. Однако в прибойной зоне флета и в прорезающих его каналах волнобой и приливно-отливные течения могут достигать такой силы, что они повреждают колонии кораллов или отрывают их от субстрата. Соответственно в этой зоне выживают и доминируют кораллы, хорошо переносящие волнобой. Это прежде всего ветвистые формы акропор, способные в зависимости от силы прибоя образовывать жизненные формы, устойчивые к волнобою и сильным течениям. В число таких кораллов входят также и ряд видов других родов ветвистых кораллов: это Pocillopora, Stylopora, Millepora. Их успешное выживание в прибойной зоне рифа обусловлено не только высокой механической прочностью их колоний и точной их ориентацией относительно направления волнового стресса (Shinn, 1963), но и высокой скоростью роста, а также их способностью размножаться обломками. Эти их свойства позволяют им быстро регенерировать после их разрушения штормовым прибоем.
Согласно выводам многих авторов, волновой стресс является главным фактором, контролирующим структуру популяций; кораллов на наветренных рифах (Geister, 1977; Jokiel, 1978; Sheppard, 1980; Dollar, 1982). Волнобой оказывает существенное влияние и на состав сообществ горгонарий и мягких кораллов, которые заселяют поверхность флета и верхние горизонты внешнего склона рифа (Morton, 1974). Во время штормов значительная их часть обрывается и выбрасывается на берег (Birkeland, 1974). На флете рифов, не испытывающих сильного волнобоя, зональность в распределении кораллов обычно выражена слабо (Loya, Slobodkin, 1971; Loya, 1972; Bouchon, 1981). Вместо обычных для внешнего края ветвистых форм кораллов их место на таких рифах занимают фавииды, Porites и Montipora (Grigg, 1983; Latypov, 1985; Potts et al., 1985; Cameron, Endean, 1985).
Влияние колебаний температуры и солености на структуру сообществ кораллов наиболее ярко выражено на осушаемом флете индо.тихоокеанских рифов. Во время осушки флета в периоды низких отливов кораллы, живущие на флете таких рифов, должны переносить пребывание на воздухе в течение 2-4 часов. При этом днем они перегреваются на солнце, ночью испытывают сильное охлаждение, а во время дождей также и распреснение (Wells, 1957; Pichon, 1964, Loea, 1972; Ditlev, 1978). При постепенном изменении солености на осушной зоне флета кораллы выдерживают колебания ее в пределах от 20 до 40% (Kinsman, 1964), но не выносят внезапного резкого распределения (Goreau, 1964). Дневная осушка рифа в ясную погоду обычно сопровождается сильным перегревом и ростом солености. Исследования показали, что стрессовые воздействия перегрева, изменений солености и избыточной освещенности на сообщества флета в периоды осушки или низких отливов взаимосвязаны и усугубляют их повреждающий эффект (Coles, Jokiel, 1978).
На осушном флете высокоширотных рифов Красного моря и Карибского бассейна основным фактором, влияющим на состав сообществ кораллов, является падение температуры воны в периода низких отливов (Loya, 1972; Scatterday, 1977; Hudson, 1981). Прорывы холодных вод с температурой ниже 16° С на рифы в этих районах вызывают гибель даже многих оппортунистических кораллов, таких, как Acropora и Pocillopora (Roberts et al., 1975). Летом на осушной зоне таких рифов кораллы выдерживают перегрев до 36°С (Kinsman, 1964). Столы суровое воздействие физического стресса выдерживают лтды некоторые наиболее стойкие опортунисты их родов: Acropora, Pocillopora, Porites, Montipora, Millepora, Diploria, Psammocora. Именно эти виды и доминируют в ассоциациях осушаемого флета (Ditlev, 1978). Существенно, что и планулы этих видов хорошо переносят условия физического стресса, оседая и выживая на осушаемом флете (Connell, 1973). Гибель кораллов может вызвать также интенсивное осаждение взвеси (Marshall, Orr, 1931). После штормов взмучивание осадка и заиление столь же губительны для кораллов, как само физическое воздействие волны или распреснения, вызываемого дождями, которыми сопровождаются шторма (Banner, 1968; Johannes, 1975). Повышенная мутность вызывает затенение дна. В условиях пониженной освещенности в мутных водах многие кораллы погибают (Rogers, 1979) или же их рост подавляется недостатком света (Dodge et al., 1974). Занос рифа осадками и плохая освещенность дна затрудняют осаждение планул и их развитие (Johannes, 1972а). В эволюционном плане стрессовое воздействие заноса кораллов осадками вызвало появление форм кораллов, устойчивых к заилению и способных активно сбрасывать с себя оседающую из толщи воды взвесь (Yonge, 1930; 1940; Lewis, Price, 1976). Такие кораллы выживают в мутных водах с низкой прозрачностью и большим содержанием взвеси в воде (Roy, Smith, 1971; Loya, 1976).
Наблюдения показали, что многие кораллы успешно выживают в очень мутных водах лагун атоллов и барьерных рифов (Wells, 1957; Roy, Smith, 1971). В их числе: Pocillopora, Galaxea, Symphillia, Fungia, Psammocora, Acropora, Porites. Эти кораллы хорошо жили в воде с высоким содержанием взвеси (Marshall, Оrr, 1931). В лагунах некоторых тихоокеанских атоллов с очень низкой прозрачностью воны (до 1 м) можно было найти рифы с высоким покрытием дна кораллами — более 30%. Так; в лагуне атолла Фаннинг, перегороженной патч-рифами, много участков с очень высокой мутностью воны и илистым дном. Тем не менее покрытие его кораллами составляло около 30%. Среди кораллов, заселявших это илистое дно, преобладали: Acropora, Montipora, Pocillopora, Stylopara. Кораллы селились пятнами, образуя на мягком дне небольшие банки. Отдельные колонии ветвистых форм селились на крупных обломках отмерших кораллов и на раковинах мертвых тридакн (Roy, Smith, 1971).
Свет как физический фактор оказывает существенное влияние на структуру сообществ кораллов лишь в самых верхних и самых нижних зонах рифа. В самых верхних горизонтах рифа имеет место угнетение кораллов избыточной освещенностью (Siebeck, 1981).
Заметное влияние недостатка света на видовую структуру сообществ кораллов сказывается только при освещенности ниже 3-5%. В этих условиях в составе сообществ начинают преобладать агерматипные вины кораллов и некоторые высокоспециализированные вины герматипных склерактиний — обитателей глубинных малоосвещенных зон рифа. Эти кораллы адаптированы к низкой освещенности и живут, не теряя своих зооксантелл, при освещенности около 1% ФАРп. Таковы некоторые вины кораллов: Leptoseris, Cycloseris, Pachyseris. В то же время в составе сообществ кораллов глубинных зон рифа при освещенности ниже 2-3% ФАРп обитают ряд видов тенелюбивых склерактиний, таких, как Leptastrea, Scolymia, Turbmaria, а также оппортунистические виды склерактиний, которые встречаются во всех зонах рифа, но способны выживать при освещенности ниже 2% ФАРп. Таковы, например, некоторые вины кораллов: Psam-mocora, Pectinia, Pocillopora. Platygyra, Seriatopora, Lobophyllia, Goniopora, Alveopora (Dinesen, 1982; Kuhlmann, 1983; Fricke, Schuhmacher, 1983; Титлянов и др., 1983; Oliver et аl., 1983; Титлянов, 1987).
Биологические факторы, определяющие развитие видовых популяций сидячего бентоса и структуру его сообществ, проявляются нз уровне межвидовых отношений выедания и конкуренции за субстрат (Paine, 1966). В ходе эволюции оба этих фактора послужили причиной выработки у них защитных приспособлений, предохраняющих их от выедания, а также появления у них средств агрессивного подавления конкурирующих с ними за пространство соседствующих донных животных. Что касается защиты от выедания, то разные группы кораллов прибегли к совершенно различной ее стратегии. Склерактинии защитили себя путем укрепления своей скелетной основы, спрятав полипы в мощные известковые крепости-кораллиты, окруженные режущими и колющими краями сети. В наибольшей степени эти защитные свойства скелета кораллитов выражены у таких кораллов, как Galaxea, Achrelia, horrescens, фунгины, а также у массивных кораллов, которые практически недоступны выеданию их рыбами. Такой же стратегии придерживаются гидрокораллы (Millepora) и два вида октокораллов Hiliopora и Tubipora, которые имеют массивную скелетную основу.
Более древние по сравнению со склерактиниями (Schmidt, 1974) октокораллы прибегли для защиты от выедания к средствам химической защиты. Содержащиеся в октокораллах в огромном количестве сложные соединения типа терпеноидов и простагландинов делают их практически несъедобными для большинства рыб, моллюсков, ежей (Bakus, 1981; Coll et аl., 1982; Gerhard, 1984).
Донные сообщества рифа ввиду необычайно высокой плотности их заселения сидячим бентосом и макрофитами характеризуются высокой напряженностью конкурентных отношений за субстрат (Connell, 1973; Jackson, 1977). Существенное влияние на их структуру оказывают конкурентные отношения между самими кораллами (Connell, 1976; Sheppard, 1980). Кораллы проявляют обычно три типа конкурентных отношений:
а) перерастание и затенение одних кораллов другими (Shinn, 1966; Porter, 1974а; Pottes, 1976; Bothwell, 1983),
б) выделение токсических метаболитов, подавляющих конкурирующие вины кораллов (Sammarco et аl., 1983),
в) межвидовая агрессия одних кораллов по отношению к другим (Glynn et аl., 1972; Lang, 1973; Dus-tan, 1975; Richardson et аl., 1979; Sheppard, 1979; Wellington, 1980; Bothwell, 1984).
В первом случае имеет место вытеснение быстрорастущими ветвистыми рамозными или крибозными кораллами растущих медленнее массивных фолиозных или ветвистых кораллов, имеющих плотные шаровидные колонии. Иногда такие ветвистые кораллы, встречая на пути роста конкурента, дают корковидные ростовые формы, которые прорастают по его поверхности и убивают его (Potts, 1976). Однако в большинстве случаев быстро растушие колонии склерактиний и горгонарий обгоняют в росте своих медленно растущих конкурентов, не приближаясь к ним на близкое расстояние и не испытывая со стороны последних агрессивной "атаки". Затеняя их, они угнетают их рост, в конце концов способны их вытеснить (Glynn, 1976; Connell, 1976; Karlson, Jackson, 1981).
Использование токсических веществ против конкурентов за субстрат широко распространено у мягких кораллов-горгонарий и альционарий, большинство из которых способны вырабатывать различные средства химической защиты и выделяют в окружающую среду токсические метаболиты (Sheppard, 1979). Используя эту способность, а также высокую скорость роста, мягкие кораллы способны перерастать и вытеснять склерактинии. Так, склероксониевая горгонария Erythropodium легко перерастает и вытесняет многие склерактиниевые кораллы и зоантарии (Karlson, 1980). Токсические метаболиты, выделяемые в воду альционарей Sinularia, подавляли в опытах рост склерактиниевых кораллов и даже вызывали их гибель (Sammarco et al., 1983).
Отношения прямой агрессии в борьбе за субстрат широко развиты среди склерактиниевых кораллов, полипы которых имеют хорошо развитый стрекательный аппарат. Оружием нападения у агрессивна их видов служат шупальца и мезентериальные нити, вооруженные нематоцистами. Приблизившись в процессе роста к соседнему кораллу-конкуренту, агрессивный коралл убивает и переваривает с их помощью своего конкурента, останавливая его дальнейший рост (Logan, 1984). Критическое расстояние, начиная с которого коралл начинает агрессию по отношению к другому кораллу, приблизившись к нему в процессе роста, специфично для каждого вида. Оно выражается величинами от 0,5-1 см у акропор и поцилопор, имеющих диаметр полипов 1-2 мм. У кораллов со средними размерами полипов (кораллитов 3-10 мм) это критическое расстояние составляет 1-4 см. Таковы Oculina и многие фавииды (Sheppard, 1981а). Многие агрессивные кораллы имеют удлиненные шупальца-"дворники". Они способны убивать полипы конкурента с помощью нематонистов, дотягиваясь до них этими щупальцами на расстояние до 3-4 cм. (Sheppard, 1981а). В числе таких видов наиболее агрессивные массивные кораллы Montastrea annularis, Diploria strigosa, Scolymia lacera, Galaxea fasricularis, (Lang, 1973; Richardson et al., 1979; Wellington, 1980; Chorneski, 1983). Шупальца-дворники вооружены усиленным книдоцилем и имеют больше крупных нематоцистов, чем обычные щупальца (Hartog, 1977; Hidaka, Yamazato, 1984). Любопытно, что число этих щупалец-органов агрессии возрастает в полипах, расположенных в той части колонии, которая в ходе ее роста приближается к потенциальному конкуренту-врагу. Если в условиях опыты выдержат такой "агрессивный" коралл вблизи другого коралла, который стоит ниже его по шкале агрессивности, то у него можно вызывать превращение части обычных щупалец в щупальца-дворники (sweeper tentacles). Такую индукцию образования щупалец-дворников удалось вызвать у коралла Agaricia agaricites, у которого в норме они вообще отсутствуют (Chornesky, 1983). Некоторые кораллы (например, Goniopora), имеют полит, которые могут вытягиваться в активном состоянии до 10-15 см и способны убить конкурента на значительном расстоянии.
Установлено, что наибольшую способность к "агрессии" в условиях рифа появляют, как правило, массивные формы склерактиний. Согласно наблюдениям Ланг (Lang; 1973), которая открыла межвидовую агрессию у склерактиний, среди них существует своеобразная иерархия по степени агрессивности (Соре, 1981). Среди атлантических кораллов наиболее агрессивные виды относятся к трем семействам: Mussidae, Meandrinidae и Faviidae. Согласно Ланг (1973), агрессивное поведение не наблюдалось при контакте двух колоний одного и того же вида, хотя это оспаривает Бусвел (Bothwell, 1984). Иерархия агрессивности отдельных видов у атлантических кораллов на рифах Ямайки, согласно Ланг, выглядит начиная с более агрессивных видов следующим образом:
Mussa anuola -> Scolymia lacera -> Isophyllia sinuosa -> Mycetophillia ferox -> Meandrina meandrites -> Mycetophyllia danaana -> Scolymia cubensis -> Isophyllastrea rigida -> Montastrea annularis -> Diploria labyrinthiformis -> D. strigosa -> D. tlivosa -> М. cavernosa -> Colpophyllia natans -> Manicina areolata -> Acropora paJmata -> А. cervicornis -> Eusmilia fastigata -> Agaricia sp. -> Helioseris cucullata -> Siderastrea -> Porites -> Stephanocoenia.
Согласно Лорану (Loran, 1984), на рифах у Бермудских островов иерархия агрессивности кораллов несколько отлична от приведенной выше. Наиболее агрессивными видами там являются Montastrea annularis, Diploria labyrinthiformis и D. strigosa. На тихоокеанских рифах в южной части Б. барьерного рифа иерархические отношения в пределах массовых семейств кораллов выглядят следующим образом: Mussidae -> Faviidae -> Acroporidae -> Pocilloporidae -> Poritidae (Bothwell, 1984). Эти данные показывают, что наименее агрессивны семейства, в состав которых входят наиболее оппортунистические видаг кораллов. В пределах этой группы иерархические отношения в значительной мере неопределенны и зависят от физиологического состояния взаимодействующих видов (Loran, 1984).
Вполне естественно, что в ходе напряженной борьбы за тверднш субстрат на перенаселенном рифе наиболее агрессивные кораллы имеют существенные преимущества, особенно в сообществах внешнего склона, где влияние факторов физического стресса на формирование сообществ ослаблено и на первый план в отношении выходят не адаптивные свойства оппортунистических видов, а конкурентноспособность агрессивных видов (Connell, 1976; Sheppard, 1980). Однако и в сообществах флета, подверженных интенсивному воздействию волнобоя, влияние агрессивных межвидовых отношений кораллов на формирование их видовой структуры прослеживается достаточно четко (Bothwell, 1984), хотя определяющее значение играет здесь физический стресс (Bradbury, Yonge, 1981; Loran, 1984).
Любопытно, что на атлантических рифах преобладают отношения агрессивного доминирования со стороны медленно растущих массивных кораллов, тогда как на индотихоокеанских рифах преобладает первый тип конкурентных отношений, когда быстро растущие ветвистые кораллы перерастают, затеняют и в конечном счете часто вытесняют медленно растущие массивные кораллы, хотя они при этом могут испытывать агрессивную "атаку" со стороны последних, если приближаются к ним на определенное расстояние (Porter, 1974; Potts, 1976; Sheppars, 1980).
Заметное влияние на структуру сообществ кораллов оказывают животные коралло-фаги-звезды акантастер, а также многие другие представители бентоса и рыб рифа. Это влияние проявляется в основном в избирательности животных в отношении поедаемых ими кораллов. Что касается влияния рыб, то они, нанося кораллам некоторый ущерб их выеданием, в значительной мере компенсируют тем, что способствуют их размножению и расселению. Питаясь кораллами, рыбы-попугаи, рыбы-хирурги, бал-листиды и кузовки дробят на кусочки веточки тонковетвистых коршглов-оппортунис-тов, таких как Acropora, Pocillopora, Stylophora, Seriatopora, Montipora. Эти обломки приживаясь на свободньк участках рифа, способствуют их расселению, облегчая им борьбу за доминирование в составе сообществ (Glynn et al., 1972; Connell, 1973; Randall, 1974; Tunniclinne, 1981; Highsmith, 1982). В то же время доказано, что на многих рифах перечисленные выше рыбы, питающиеся кораллами, отдают предпочтение определенным их видам, таким, как например, Pocillopora damicornis. Выедание этого коралла рыбами в сообществах склона может идти столь интенсивно, что этот вид вообще из них исчезает и встречается лишь в сообществах флета и края рифа, где прибой мешает его выеданию. Перенесение колоний поциллопоры с флета на склон на глубину 15-30 м вызывало быстрое их поедание рыбами (Neudecker, 1979).
Те же рыбы, а также многие рыбы-фитофаги, которые выедают водоросли и перифитон на скальный поверхностях рифа и на отмерших кораллах, очищают тем самым место для осаждения гшанул (Vine, 1974; Brikeland, 1977; Sutton, 1983). Влияние рыб-кораллофагов, равно как и многих представителей бентоса, питающихся кораллами, на структуру их сообществ обусловлено избирательностью кораллофагов в отношении поедаемых ими видов кораллов и их возрастньк групп. Чаще всего ими повреждаются молодые колонии ветвистьк и гшастинчатьк кораллов, растущие вблизи крупньк колоний кораллов, которые служат убежищем рыбам и крабам, поедающим кораллы (Grassle, 1973).
Существенное влияние на формирование и структуру сообществ кораллов оказывают ежи Diadema (Sammarco et al., 1974; Sammarco, 1980, 1982). Их присутствие в умеренном количестве (1-4 эзк/м2) сопсобствует осаждению гшанул на участках скального флета, которые они освобождают от водорослей. Тем сампгм способствуют увеличению длотности заселения субстрата кораллами. В случае же очень высокой их концентрации на флете, которая может достигать 60 экз/м2, ежи повреждают спат кораллов и их молоднге колонии, подавляя таким образом их развитие (Bak, Van Eys, 1975). Другие представители бентоса, поедающие кораллы, такие, как полихета Hermodyce carunculata и гастропода Carallophila, проявляют пищевую избирательность, предпочитая одни кораллы другим. Тем самым они также могут влиять на структуру их сообществ (Rylaarsdam, 1983). В прибойной зоне особенно заметное влияние на сообщества кораллов оказывают сверлящие губки Cliona и моллюски Lithophaga, Fungioclava, Siphonodyotion (Kleemann, 1973, 1980; Hein, Risk, 1975; Wilkinson, 1983). Они ослабляют прочность их известкового скелета, что способствует их разрушению под действием волнобоя.
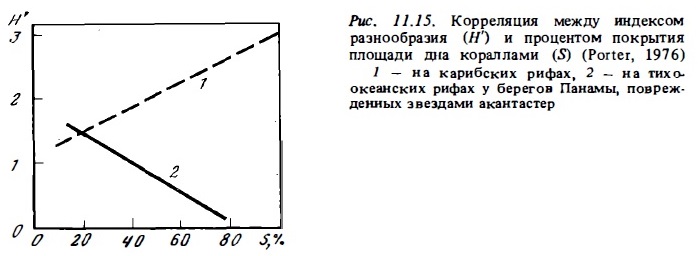
Звезда акантастер поедают прежде всего доминирующие оппортунистические виды кораллов. Вполне естественно, что после их нападения структура сообщества кораллов меняется. Их видовое разнообразие растет, хотя общая их численность (процент покрытия) снижается. Такое явление быто описано Портером (Porter, 1972) на тихоокеанских рифах у Панамы (рис. 11.15). В данном случае рост показателя видового разнообразия на рифе, подвергшемся нападению звезд, быт связан с тем, что они истребили преимущественно популяцию доминирующего здесь коралла Pocillopora (Porter, 1974а). На других рифах звезда выедали в основном акропор, предпочитая более молодые колонии этих, а также некоторый других оппортунистических видов фунгид и поритид (Done, 1985).