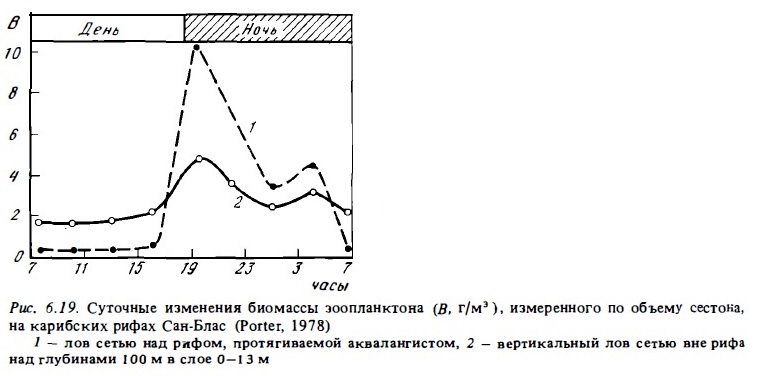
Применение ловушек для учета количества демерсального зоопланктона, мигрирующего ночью со дна в толщу воды, показывает, что масса мигрирующего планктона достаточно велика и может достигать сотни граммов на 1 м2 (табл. 6.17), или 1— 5тыс./м2 за ночь (рис. 6.19, табл. 6.17). Эти данные подтверждают результаты ночных сетных ловов, выявивших в водах над рифами необычайно богатый зоопланктон (см. табл. 6.23). При этом наиболее богатым оказался ночной зоопланктон мелководный участков рифа в зоне флета, патч-рифов, живых кораллов, скоплений обломочного материала, т.е. именно там, где, по прежним представлениям (Johannes et al., 1970; Glynn, 1973а), его биомасса должна быть наименьшей, поскольку здесь он должен был бы наиболее интенсивно выедаться донными хищниками, прежде всего кораллами. Такое выедание, безусловно, имеет место (Johannes, Gerber, 1974, см. табл. 6.18). Однако период интенсивного выедания по времени не должен были продолжительным, поскольку оно происходит главным образом при выходе демерсальньк форм в толщу воды и при их возвращении, т .е. в течение относительно коротких промежутков времени (Porter, 1974). По оценкам авторов, определявших выедание зоопланктона сообществом рифа (Glynn, 1973а; Johannes, Gerber, 1974), оно должно определяться максимум первыми десятками миллиграммов на кубический метр, или около 100-150 экз./м3, тогда как ночная миграция зоопланктона из донных субстратов достигает нескольких граммов на квадратный метр по биомассе и тысяч экземпляров на кубический метр по численности (см. табл. 6.16, 6.17) .
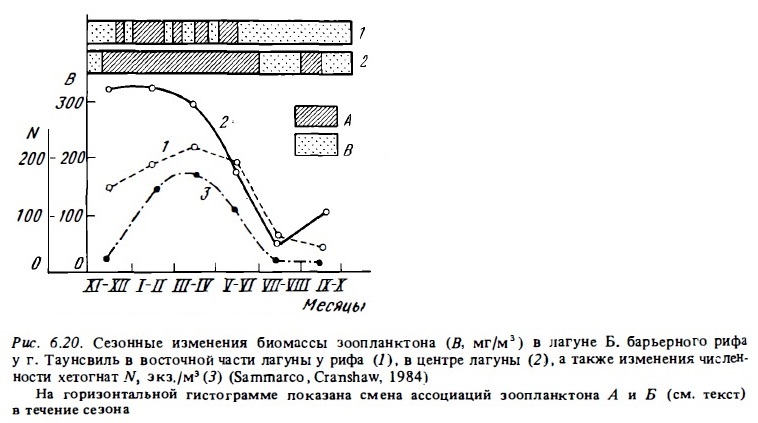
Очень скудны данные о сезонной динамике биомассы зоопланктона в рифовых водах. Судя по результатам имеющихся наблюдений, она может испытывать значительные колебания в разные сезоны года, как это имеет место с биомассой фитопланктона. Например, в лагуне Б. барьерного рифа в течение года происходят закономерные изменения состава популяций зоопланктона (рис. 6.20). Его биомасса (по данным дневных ловов) при этом меняется от максимальной (290-300 мг/м3) зимой до минимальной (50-60 мг/м3) летом. Отмеченный здесь максимум зоопланктона в зимний период непосредственно связан с возрастанием зимой более чем на порядок объема речного стока, с которым в лагуну поставляются биогенные элементы (Sammarco, Crenshaw, 1984). Аналогичное явление описано для зоопланктона рифов Новой Каледонии (Binet, 1984).