В лагунах атоллов с низкой первичной продукцией (атоллы о-ва Лау и Маршало-вых островов, островов Туамоту) нижний предел (2-4 мг С/м3 в сутки) близок к ее величинам в омывающих их олиготрофных тропических водах пассатных течений, где она составляет обычно 1-5 мг С/м3 в сутки (Сорокин, 1971а). Однако и в таких атоллах в водах их лагун возможны сезонные вспышки развития фитопланктона, которые могут достигать силы "цветения" (Sournia, Ricard, 1976а). Как видно из данных табл. 6.7, в лагунах атоллов, даже в тех из них, которые омываются олиготрофными тропическими водами с крайне низким уровнем развития фитопланктона, первичная продукция чаще всего соответствует среднему, а иногда и верхнему уровню продуктивности фитопланктона в мезотрофных морских бассейнах (15-50 мг С/м3 в сутки). В лагуне крупного кольцевого рифа Каргадос-Карахос, расположенного в стрежне очень малопродуктивных вод Южного пассатного течения Индийского океана, первична продукция достигала 250 мг С/м3 в сутки при биомассе водорослей (преимущественно диатомей) более 5,2 г/м3 (табл. 6.10). Высокая продукция (70100 мг С/м3 в сутки) была зарегистрирована в закрытой лагуне атолла Фаннинг (острова Лау) в период развития там динофлагеллят и кокковидных синезеленых водорослей (Gordon et al., 1971). Этот атолл также располагается в олиготрофных водах пассатного течения. Таким образом, даже в лагунах отдельных кольцевых рифов, расположенных в потоке олиготрофных вод, могут существовать достаточно высокопродуктивные сообщества фитопланктона, продукция и биомасса которых на 1-2 порядка выше, чем в омывающих их водах океана.
Судя по результатам имеющихся немногочисленных измерений, концентрации фотосинтетически активного хлоофилла "а" в воде лагун кольцевых рифов открытого океана в среднем в 5-10 раз выше, чем в окружающих водах океана. По данным единичных измерений, она варьировала в лагунах разных рифов в пределах от 0,2 до 1 мг/м3. В лагуне атолла Такапото, по результатам длительных наблюдений (Souгnia, Ricard, 1976а) , концентрация хлорфилла менялась в пределах от 0,26 до 0,77 мг/м3, причем она заметно варьировала и в разных частях лагуны в зависимости от интенсивности поступления в них вод океана во время приливов .
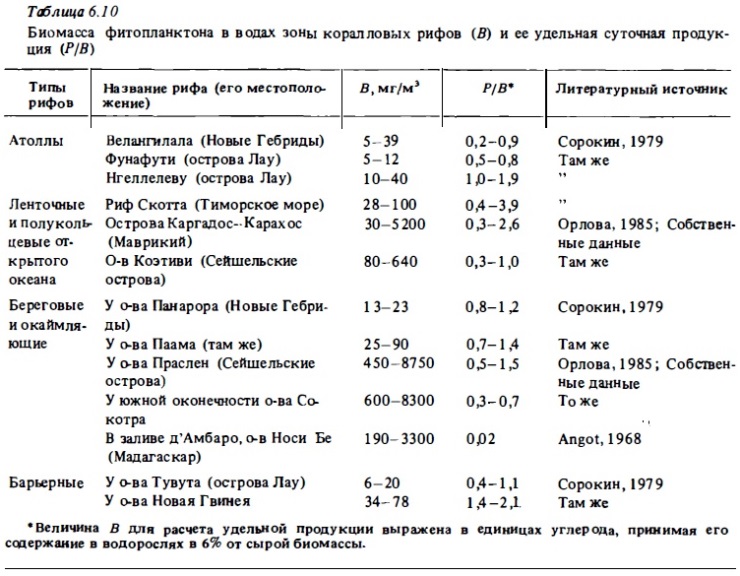
Существенный вклад в изучение фитопланктона и его продукции дали сезонные наблюдения, выполненные в водах барьерных и береговых рифов вблизи крупных островов, где были основаны биологические станции доя изучения жизни рифов и прилегающей неритической зоны океана. В числе таких хорошо изученных рифов следует упомянуть рифы у о-ва Носи Бе и у Тулеара (Мадагаскар), Гавайский риф у о-ва Оаху, Б. Барьерный риф Австралии, рифы Вапрао, Тиаура и Фааа на островах Таити и Муреа, рифы центрального Вьетнама вблизи г. Нячанг. Результаты измерения первичной продукции и биомассы фитопланктона барьерных и береговых рифов обобщены в табл. 6.8, 6.9, 6.10: Представленные в них данные очень сильно варьируют почти для всех изученных рифов. Они отражают закономерные измерения первичной продукции в разных зонах одного и того же рифа. При этом наименьшие величины ее (так же как и наименьшие величины биомассы фитопланктона: рис. 6.10) соответствуют мелководным зонам рифа, где вода находится в интенсивном контакте с донным населением и альгофлорой рифа. В этих зонах (флет, патч-рифы, литораль) фитопланктон, как правило, бывает обедненным и его продукция здесь резко падает. Максимальные величины продукции и биомассы фитопланктона, представленные в указанных таблицах, на каждом рифе соответствуют зонам лагуны или внешнего склона окаймляющего или берегового рифа с глубинами более 15-20 м, где вода находится вне прямого контакта с донным населением рифа. Величины первичной продукции в этих зонах, как правило, бывают значительно выше, чем в водах прилегающей акватории открытого моря. В среднем они выше, чем в воде лагун атоллов и кольцевых рифов открытого океана.
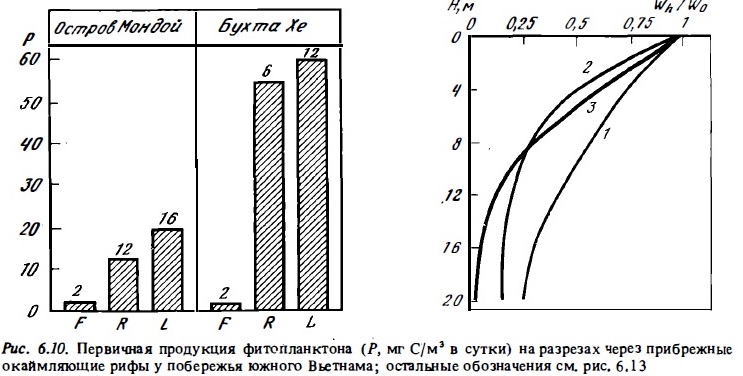
Высокие величины первичной продукции, достигающие уровня евтрофных вод, были отмечены в воде ряда береговых и барьерных рифов Индийского океана, таких, как рифы у Мадагаскара, у Сейшельских островов и у о-ва Сокотра, а также в лагуне Б. барьерного рифа в периодах "цветения" синезеленых водорослей (см. рис. 6.9). Суточная продукция достигала здесь 50-300 мг С/м3 в сутки в верхнем слое воды, или 0,7-2,8 г С/м3. При этом в водах барьерных рифов островов, омываемых олиготрофными водами океана (Танги, Оаху, Лизард), первичная продукция в лагунах может достигать 50-60 мг С/м2, или 0,4-0,7 г углерода в 1 м2 в сутки, что соответствует верхнему уровню продуктивности мезотрофных вод. В рифовых зонах побережья Центрального Вьетнама и его островов в Южно-Китайском море (острова Тху) и в Сиамском заливе (острова Намзу) (см. табл. 6.8, рис. 6.10) величины первичной продукции фитопланктона были высокими, несмотря на то что наблюдения производились в период сезонного минимума фитопланктона (Сорокин и др., 1982). Наибольшие величины первичной продукции, как правило, наблюдались в водах у внешнего края рифа на расстоянии 10-100 м от него над глубинами 5-15 м (см. рис. 6.10). Она обычно превышала здесь 30-40 мг С/м3 в сутки. На отдельных разрезах в этой зоне в 1984 г. была зарегистрирована первичная продукция выше 100 мг С/м3 в сутки.
Сведения о биомассе фитопланктона береговых и барьерных рифов пока еще весьма отрывочны. Наиболее подробные измерения биомассы фитопланктона в течение сезона были выполнены Анго (Angot, 1968) у о-ва Носи Бе (Мадагаскар). В .обобщенном виде результаты этих измерений приводятся на рис. 6.9. В табл. 6.10 представлены имеющиеся в литературе данные о биомассе фитопланктона ряда крупных рифов Индийского и Тихого океана. Как видно из таблицы, биомасса фитопланктона в водах барьерных и береговых рифов может достигать 3-8 г/м3. Самые низкие виличины ее были отмечены в воде береговых и барьерных рифов центральной части Тихого океана, находившихся в угнетенном состоянии (рифы у островов Панарора и Паама, Новые Гебридах, риф у о-ва Тувута, острова Лау).
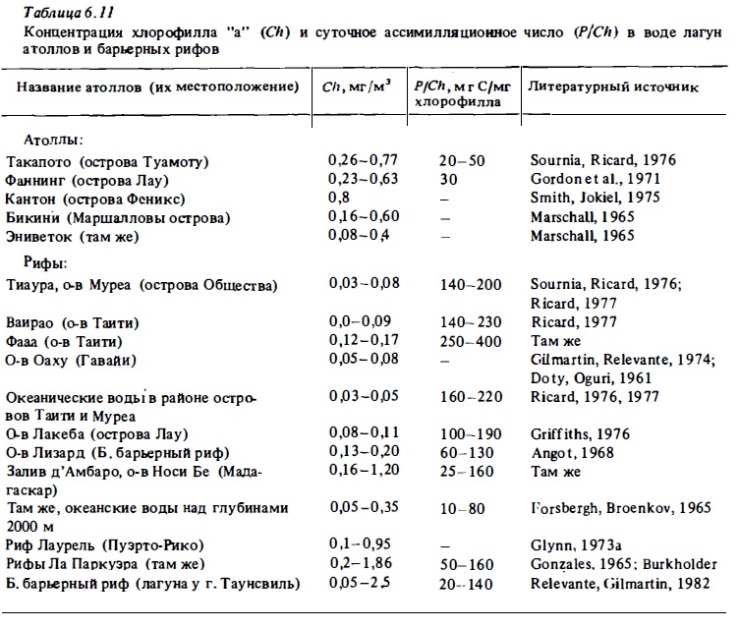
Столь же ограниченны сведения о содержании хлорофилла "а" в водах барьерных и береговых рифов (табл. 6.11). Наиболее подробные ее измерения были выполнены в водах лагун островов Муреа и Таити французскими исследователям (Ricard, 1976; Sournia, Ricard, 1976). Эти измерения показали, что концентрация хлорофилла в них в разные сезоны года варьирует в пределах 0,03-0,17 мг/м3. В лагуне Гавайского рифа у остра Оаху, а также в лагуне Б. барьерного рифа зимой концентрация хлорофилла была близка к таковой в мезотрофных морских водах: 0,1—0,3 мг/м3. В периода осеннолетних максимумов развития фитопланктона в лагуне Б. барьерного рифа, а также в водах рифа у о-ва Носи Бе (Мадагаскар) концентрация хлорофилла в воде была выше на 1 мг/м3, что близко к его содержанию в евтрофных водах (Angot, 1968). Установлено влияние взмучивания всего слоя донных осадков, обогащенного микробентосом, в зоне волнобоя на концентрацию хлорофилла в водах рифа (Walker, 1981).
Как уже отмечалось выше, величины первичной продукции и биомассы фитопланктона сильно варьируют в разных зонах одного и того же рифа. В большинстве случаев самые низкие величины биомассы и продукции фитопланктона характерны для мелководных зон рифа и особенно для риф-флета, где вода, проходя над рифом во время приливов и отливов, находится в наибольшем контакте с донным населением рифа. Причину убыли фитопланктона на мелководных участках флета следует видеть в его выедании донными фильтраторами рифа (Glynn, 1973а). К тому же клетки днатомей, таких, как Nitzschia delicatissima или ветвистные клетки Chaetoceros, могут травмироваться в турбулентных водах рифа, особенно при столкновениях с твердой поверхностью рифа и с частицами взвеси. В водах же приглубых частей лагуны или за внешним краем рифа, где контакт вод с рифом уменьшается, происходит интенсификация развития фитопланктона за счет отдачи части биогенов в оду донными сообществами рифа.
По количественным показателям его биомассы и продукции, по концентрации хлорофилла фитопланктон в зоне рифа богаче, чем в окружающих водах открытого океана (Michel, 1969; Сорокин, 1971; Ricard, 1976; Sournia, Ricard, 1976а). На рис.6.3 была представлена типичная картина изменения первичной продукции фитопланктона на разрезах от рифа Херон (Б. барьерный риф) в открытый океан. Уже на расстоянии 6-9 км она убывает более чем на порядок величин.
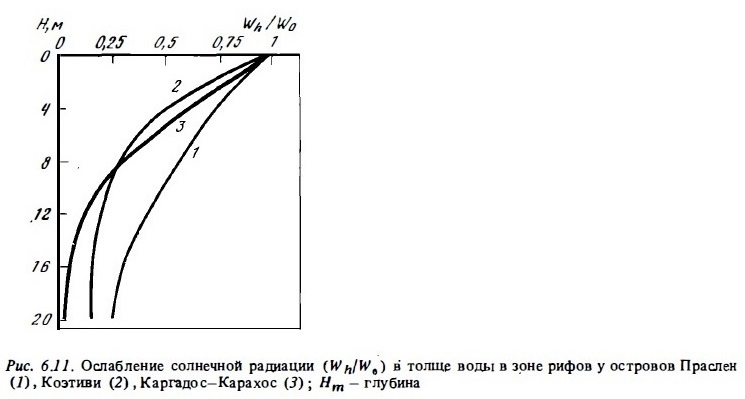
Прозрачность вод по белому диску в приглубых участках рифов обычно достаточно велика и составляет в среднем 10-15 м, хотя в отдельных случаях она может быть меньше (5 м на атолле Кантон) и больше 30 м (атолл Маджуро). Таким образом, фотосинтез фитопланктона практически не лимитируется светом во всей их толще. На рис. 6.11 приводятся типичные кривые ослабления падающей радиации в водах некоторых рифов Индийского океана со средней прозрачностью (10-18). Как видно из этого рисунка, фитопланктон может испытать некоторый недостаток света лишь у самого дна лагуны.
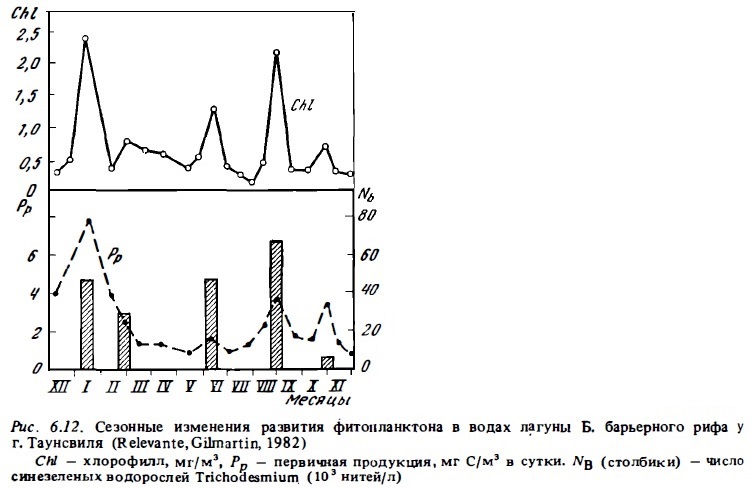
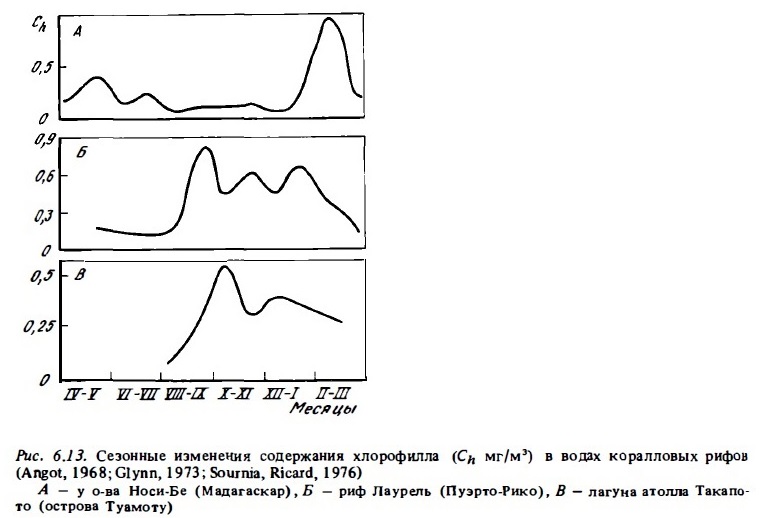
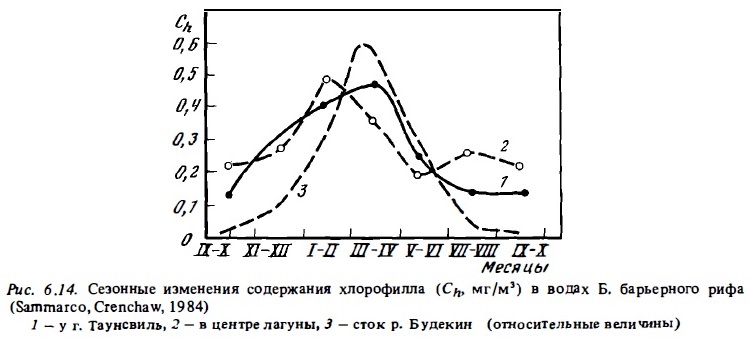
Примеры сезонной изменчивости фитопланктона представлена на рис. 6.9,6.12,6.13. Они показывают, что в разные сезоны года не только меняется его состав (см.6.2.1), но происходят изменения его биомассы и функциональной активности. Колебания величин первичной продукции фитопланктона и концентрации хлорофилла на рифах в течение сезона достигали порядка величин (Angot, 1968; Sournia, 1972). При этом даже в лагуне атолла Такапото, расположенного в районе олиготрофных вод, концентрация хлорофилла в период сезонного максимума достигала уровня, характерного для мезотрофных вод (Sournia, Ricard, 1976). В водах барьерного и берегового рифов его концентрация бьта близка к 1 мг/м3, что соответствует его содержанию в евтрофных водах, а биомасса фитопланктона в зоне рифов у о-ва Носи Бе превышала 3 г/м3. Значительные сезонные колебания концентрации хлорофилла в воде (от 0,13 до 0,5-2,5 мг/м3) были отмечены в лагуне Б. барьерного рифа Австралии у г. Таунсвиль (Sammarco, Crenshaw, 1984). Максимум был отмечен в летне-осенние месяцы с декабря по апрель. Он совпадал с периодом дождей и увеличением поверхности стока (рис. 6.14). В этот период в водах лагуны отмечаюсь интенсивное развитие фитопланктона, значительную долю которого составляли синезеленые водоросли Trichodcsmium (Relevante et al., 1982). Стимулирующее влияние речного стока на развитие фитопланктона отмечено и для вод рифов у Новой Каледонии (Binet, 1984). Представленные данные указывают на неосновательность тезиса (Parsons et al., 1977) о том, что планктонные сообщества тропических вод в отличие от умеренных находятся в состоянии некоего гомеостаза и не испытывают существенных сезонных изменений. Такие представления были, по-видимому, следствием недостатка соответствующих сезонных циклов наблюдений в тропических водах (Sournia, 1969). Причина сезонных максимумов в развитии фитопланктона прибрежных тропических вод состоит прежде всего в изменении циркуляции вод на прилегающей к рифам акватории океана, которые имеют годовую периодичность. Причем эти изменения даже в тропической зоне чаще всего имеют место в осенний и весенний период. Не случайно поэтому и сезонные максимумы развития фитопланктона отмечены осенью или весной.
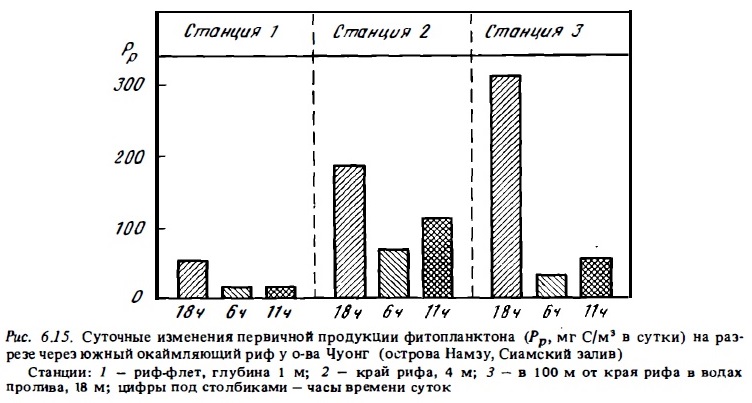
В ходе изучения развития фитопланктона в водах рифов установлено наличие значительных суточных колебаний состава и численности его сообществ, а также его продуктивности (Milliman, Mahnken, 1972; Sournia, Ricard, 1976; Сорокин, 1979). Такие колебания имеют место и в сообществах фитопланктона открытого океана. Они вызываются там суточной периодичностью роста фитопланктона и интенсификацией выедания водорослей ночью зоопланктоном, мигрирующим из нижележащих слоев воды в эвфотическую зону. Те же причины действуют и в прибрежных водах в зоне коралловых рифов, вызывая суточную периодичность развития фитопланктона. Но в этих водах на них накладывается еще влияние суточного приливного цикла. Как показали результаты суточных наблюдений на рифах у островов Намзу, под влиянием выедания мигрирующего ночью в толщу вода фито1шанктона зоопланктоном рифа суточная его продукция ранним утром снижается на порядок величин. Однако уже к 11ч популяция фитопланктона в значительной степени восстанавливается, особенно у края рифа, где фитопланктон интенсивно размножается (рис. 6.15).
Каковы же причины возрастания биомассы и продукции фитотанктона в рифовых водах по сравнению с окружающими водами открытого океана? Вопрос этот достаточно сложен, особенно ввиду того, что такое возрастете имеет место при очень низких абсолютных концентрациях биогенов, мало отличающихся от таковых в открытом океане, а также в условиях необычайно интенсивной эксплуатации сообществ фитопланктона фильтрирующей фауной рифа. Высказанные по данному поводу мнения весьма противоречивы (Sournia, Ricard, 1976; Sournia, 1976б; Salvat 1982). Большая часть авторов в качестве основной причины указывают на так называемый островной эффект ("island mass effect"). Этот эффект заключается в обогащении биогенными элементами вод прибрежной зоны у‘ островов за счет взаимодействия их массы с набегающим потоком океанских течений, в результате чего возникает подъем глубинных вод к поверхности. Эффект обогащения неритической зоны у островов, расположенных в стрежне пассатных течений, таких, как, например, Маркизекие острова (Sournia, 1976в). Б. барьерный риф (Gilmartin, Relevante, 1974), Гавайи (Doty, Capurro, 1959), Сейшельские острова (наши наблюдения), несомненно, имеет место. Под его воздействием первичная продукция в неритической зоне у таких островов бывает достаточно высокой, что влияет на уровень продуктивности и самих рифовых вод. Однако и в этих условиях продукция и биомасса фитопланктона в водах лагун рифов обычно выше, чем в прииегающих к ним и находящихся с ними в состоянии интенсивного обмена водах неритической зоны (Marshall, 1965; Michel, 1969; Sournia, Ricard, 1976, 1976а; Thompson, Golding, 1982). В свете этих данных можно сделать вывод о том, что основная причина повышенной продуктивности фитопланктона в рифовых водах состоит в постоянной отдаче в толщу воды донными и планктонными гетеротрофами биогенных элементов (прежде всего азота, фосфора), которые образуются в результате процессов деструкции органического вещества, протекающих на рифах с большой силой. В то же время ввиду очень высокой скорости оборота биогенов в толще воды рифов далеко не всегда можно зарегистрировать прирост абсолютный концентраций солевых биогенов в водах в зоне рифа по сравнению с прилегающими водами океана (рис. 5.1.).