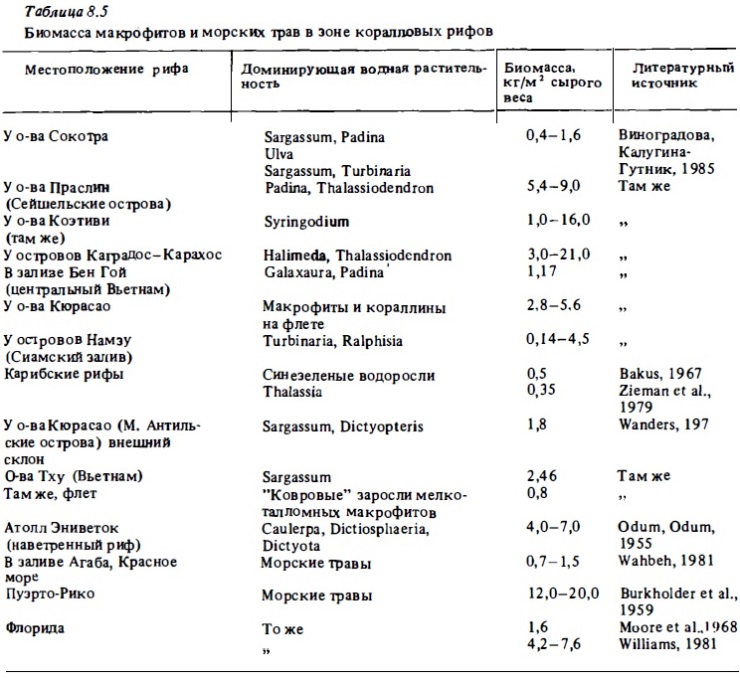
Среднее содержание хлорофилла "а" в растительных ассоциациях дна варьирует от 100 до 250 мг/м2 при их биомассе 1,5-4 кг/м2 в сыром весе. Таким образом, на 1 мг хлорофилла в среднем приходится около 15 г сырой биомассы. При этом содержание хлорофилла в расчете на площадь дна в ассоциациях с доминированием кораллин и талломньк макрофитов имеет близкие значения (Wanders, 1976).
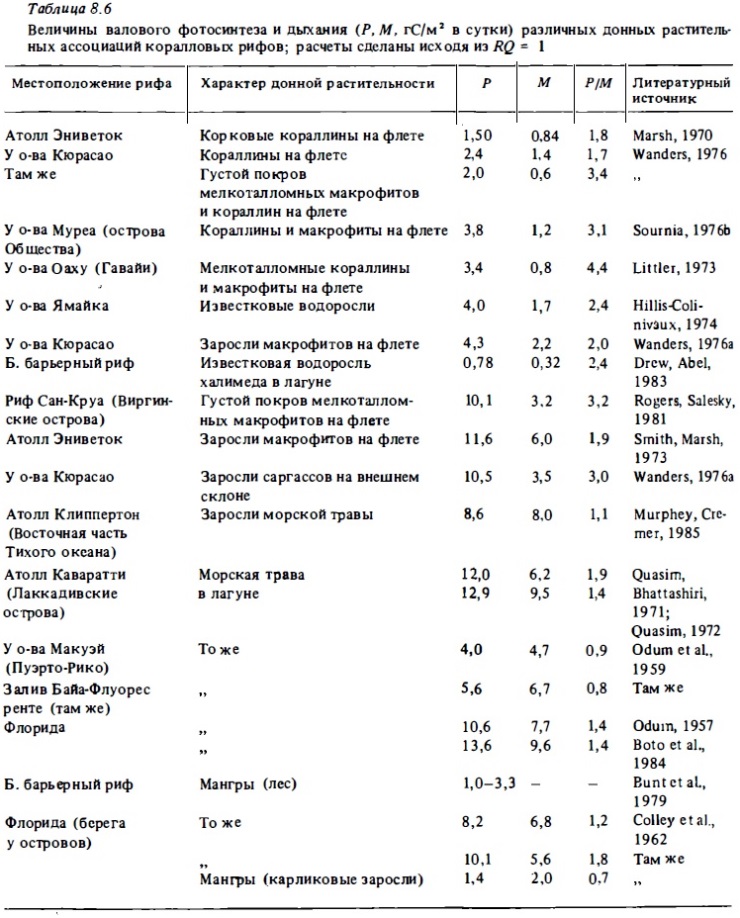
Многочисленные измерения первичной продукции фотосинтеза донной растительности кораллового рифа были выполнены как для отдельных видов растений (Quasim, Bhattathiri, 1971; Wanders, 1976; Титлянов и др., 1983), так и для их ассоциаций, обитающих в разных зонах рифа (Marsh, 1974; Wanders, 1976a; Murphey, Cremer, 1983, и др.). Определения производились с применением метода С1 4, методов регистрации колебаний кислорода in situ, а также в замкнутых и проточных сосудах. Результаты измерений интенсивности фотосинтеза и дыхания у некоторых массовых видов корковых кораллин, обитающих на поверхности флета и на его внешнем склоне, суммированы в табл. 8.6. Их валовой фотосинтез выражался величинами 1,0-2,3 г С/м2 в сутки, что соответствует средней интенсивности их фотосинтеза в середине дня 10-20 мкг С, или 0,03-0,06 мкг О2/см2 в час (Marsh, 1970; Littler, 1973а; Wanders, 1976). Фотосинтез кораллин в суточном балансе в 2-4 раза выше дыхания, интенсивность которого составляет у них 40-100 мкг С/см2 в сутки. Соответственно их чистая продукция выражается величинами 40-150 мкг С/см2 в сутки. При такой величине их чистой продукции скорость линейного роста корковых кораллин оказывается весьма значительной: 10-20 мм в год, или до 25 км в сутки (Agegian, 1981), что близко к скорости роста массовых кораллов.

Кривая суточного хода фотосинтеза у кораллин (Wanders, 1976) оказалась сходной с таковой у кораллов (см. рис. 12.13). Полученные в соответствии с этими кривыми соотношения между часовой интенсивностью фотосинтеза, измеренной в пределах от 10 до 16 ч, и суточной ее величиной, по нашим измерениям, для перифитона рифа составляют 9-10,5. По данным Уандерса (Wanders, 1976), для кораллин и сообществ макрофитов флета оно колеблется в тех же пределах: 8,7-10,7, составляя в среднем около 9,6. Мы пользовались такой величиной пересчетного коэффициента для расчета суточных величин фотосинтеза при составлении обобщающих таблиц, характеризующих суточную продукцию растительных ассоциаций рифа.
Величина подводной освещенности, при которой валовой фотосинтез уравновешивается дыханием, а чистая продукция равна нулю (компенсационная точка), для корковых кораллин флета лежит в пределах 0,9-2,5 к·лк. Это соответствует 1-3% проникающей фотосинтетически активной солнечной радиации (ФАР) в середине дня. На большинстве океанских рифов с прозрачностью вод 15-40 м по белому диску такая освещенность будет на глубинах 40-80 м. Так, у кораллин, обитающих на внешнем склоне рифов Маршалловых островов, компенсационная точка фотосинтеза отмечается на глубине около 80 м (Marsh, 1970). Световое насыщение фотосинтеза у кораллин наступает при освещенности 7-10,5 к·лк (Marsh, 1970; Wanders, 1976, рис. 8.4). В целом эта величина близка к аналогичному уровню у талломньх макрофитов: 9-10 к·лк (рис. 8.5). Таким образом, массовые виды кораллин, обитающих на флете, адаптированы к достаточно высокой солнечной радиации. В этом отношении они мало отличаются от других сопутствующих водных растений.
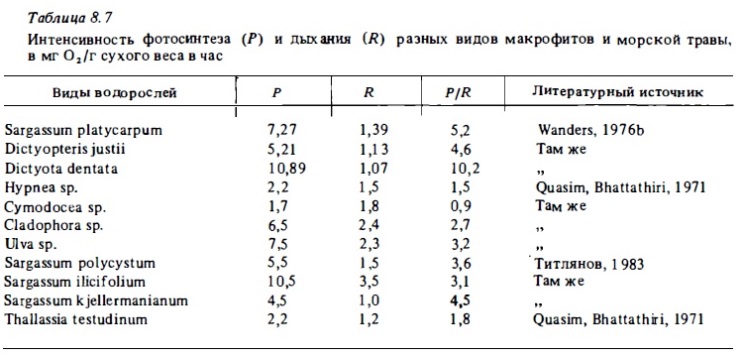
Ассоциации талломных макрофитов характеризуются весьма высоким уровнем первичной продукции — до 10 г С/м2 в сутки (табл. 8.6). Это обусловлено необычайно высокой интенсивностью валового фотосинтеза водорослей в условиях рифа. Последняя для большинства их видов близка к 2-4 мг О2/г в час, а у некоторых превышает 10 мг О2/г (табл. 8.7). Наименьшие величины интенсивности фотосинтеза в расчете на сухой вес характерны для известковых водорослей (Halimeda, Penicillus) ввиду нагруженности их талломов инертным скелетным известняком. В расчете же на поверхности таллома интенсивность фотосинтеза халимеды вдвое выше, чем у кораллин. Суточная величина коэффициента Р/В у халимеды близка к 0,07 (Drew, Abel, 1983). Зеленая известковая водоросль Penicillus дает 4—8 генераций в год при средней продолжительности жизни таллома около 2 мес (Vadas et al ., 1980).
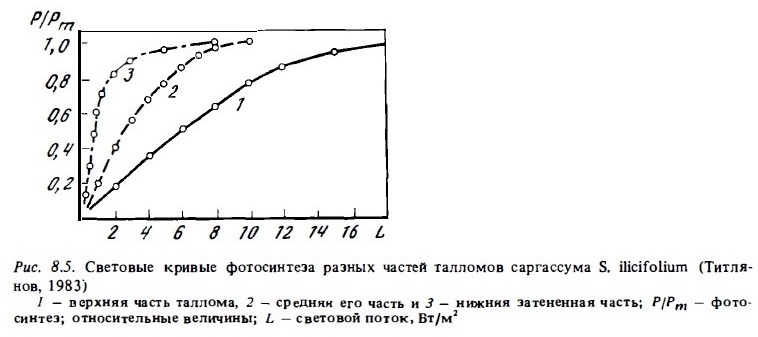
Характер световых кривых макрофитов сильно варьирует даже в пределах одного и того же вида в зависимости от времени суток и интенсивности наружной освещенности в момент взятия таллома для измерений (Титлянов и др., 1983). Оптимум освещенности, соответствующий уровню светового насыщения фотосинтеза, варьирует в среднем в пределах от 5 до 10% энергии проникающей фотосинтетической радиации (ФАР). Он составляет от 8 до 30 Вт/м2 при уровне энергии проникающей ФАР у поверхности 250-350 Вт/м2. Величина оптимума освещенности (точка светового насыщения фотосинтеза) может меняться в пределах от порядка величин в опытах с нижними и верхними частями таллома одной и той же водоросли, если его нижняя часть постоянно находится в условиях затенения (см. рис. 8.5). Эти данные говорят о способности фотосинтетического аппарата морских макрофитов к быстрой перестройке и адаптации к фактическим условиям освещенности, что в значительной мере и определяет высокую автотрофную продукцию их ассоциаций в условиях рифа.
В числе основных механизмов световой адаптации водорослей или частей их талломов, находящихся в условиях затемнения, — повышение концентрации в них хлорофилла и изменение формы таллома. Так, у саргассов нижняя затененная часть таллома содержит в 1,5-3 раза больше фотосинтетичесКих пигментов, чем верхние хорошо освещенные его части (Титлянов, 1983). Примером изменения формы талломов макрофитов, обитающих при пониженной освещенности, может быть зеленая водоросль Caulerpa, обитающая на внешнем склоне рифа. На глубинах более 10 м талломы этих водорослей имеют длинные тонкие веточки, тогда как у растений, живущих выше 10 м, ветви таллома короткие и массивные (Jaubert, 1981).
Сведения о продуктивности и метаболизме растительных ассоциаций рифа суммированы в табл. 8.6. Для участков флета, занятых кораллинами в смеси с покровом мелкотрофных макрофитов или нитчатых зеленых и синезеленых водорослей, валовая продукция фотосинтеза имеет обычно средние значения 3,5-5 г С/м2 в сутки при отношении валовый фотосинтез-дыхание (P/R), близком к 2-4. Таким образом, чистая автотрофная продукция таких ассоциаций должна выражаться величинами 2,5-4 г С/м2 в сутки. Суточные величины Р/В-коэффициентов в них близки к 0,02-0,03. В зарослях халимедаг валовая продукция фотосинтеза близка к 1-4 г С/м2. В зарослях макрофитов с относительно крупными талломами (Sargassum, Ulva, Caulerpa, Cystoseira), а также в плотных зарослях мелкоталломных водорослей без кораллин продукция фотосинтеза достигает максимума: 10-11,6 г С/м2 в сутки при отношении P/R 2-4 и чистой продукции 4-7 г С/м2 в сутки.
Сообщества морских трав характеризуются наибольшим уровнем валового фотосинтеза в расчете на единицу площади дна не только среди других биотопов рифа, но и вообще среди любых других природных растительных ассоциаций, исключая лишь тропически влажный лес (Odum, 1957; Zieman et аl., 1979). Величина валового фотосинтеза может достигать в них 12-20 г С/м2 в сутки (Quasim, Bhattashiri, 1971; Murphey, Kremer, 1983). Средние величины валового фотосинтеза в зарослях морских трав в зависимости от их густоты варьируют в пределах 5-10 г С/м2 в сутки при соотношении P/R 1-1,5. Таким образом, чистая продукция в ассоциациях морских трав оказывается относительно небольшой: 1-4 г С/м2. Часто она бывает даже отрицательной ввиду интенсивного дыхания как самих растений, так и находящихся в пределах их ассоциаций гетеротрофов: микробентоса, эпифитов и донной микрофлоры (см. табл. 8.6). Наряду с определениями фотосинтеза чистую продукцию морских трав определяли по приросту биомассы листьев (Greenway, 1974; Zieman, 1975). Полученные при этом величины составляли 1,2-2,4 г С/м2 в сутки. Биогенное питание морских трав осуществляется как с помощью корневой системы, так и через их листовую поверхность (Zieman, 1975; Whiams, 1981). Значительную долю необходимого азота поставляют им обитающие на их листьях азотфиксирующие эпифитные синезеленые водоросли (Capone et аl., 1979).
Оценки первичной продукции мангров, полученные разными методами, дали величины, которые варьировали в пределах 3-83 кг С/гектар в сутки (0,3-10,1 г С/м2). Средняя ее величина была близка к 20-30 кг С/га в сутки (см. табл. 8.6). В расчете на листовую поверхность часовая продукция фотосинтеза мангров несколько ниже 172 таковой у водорослей и составляет 130 мкг С/см2 в час (Christiensen, 1978; Bunt et al., 1979). Вынос органического вещества из зарослей мангров в прилегающие к ним донные биотопы рифа оцениваются величиной, близкой к 1 г С/м2 в сутки при первичной продукции сообщества зарослей около 8 г С/м2 в сутки (Colley et al., 1962).
Большую роль в экологии и продуктивности растительных ассоциаций рифов играют водоросли-эпифиты, растущие на поверхности талломов макрофитов, на листьях морских трав и на воздушных корнях мангров. Установлено, что между растением-хозяином и поселившимися на нем эпифитами происходит обмен продуктами фотосинтеза (Penhale, Smith, 1977), а также биогенами (Penhale, Thауеr, 1980). Эпифиты-азотфик-саторы, представленные синезелеными водорослями и азотбактером, поставляют растению-хозяину необходимый ему азот, получая от него энергетический материал в виде продуктов его фотосинтеза. Синезелешие водоросли-азотфиксаторы, обитающие на саргассах и морских травах как эпифиты, могут обеспечивать поставки растению-хозяину до 40% необходимого ему азота (Hanson, 1977; Capone et аl., 1979; Penhale, Capone, 1981). Таким образом, отношения между эпифитами и их хозяевами сходны с симбиозом.
По средним оценкам, продукции биотопов флета с доминированием кораллин приблизительно в 2 раза ниже продукции фотосинтеза на участках дна с доминированием кораллов, которая в среднем близка к 5—8 г С/м2 в сутки. Продукция же донных биотопов с доминированием морских трав и макрофитов не только соизмерима, но в большинстве случаев даже выше последней.
Представленные материалы доказывают важное функциональное значение донной растительности в экосистеме рифа. Оно состоит прежде всего в том, что донные растения рифа, такие, как нитчатки, талломные макрофиты, кораллины, морские травы, производят значительную долю общей автотрофной продукции органического вещества. Их вклад в общую продукцию фотосинтеза различен на разных рифах. Но в среднем его можно оценить в 30—50%, а на "водорослевых" рифах со слабым развитием на них живых кораллов — до 70—80%. Весьма существенно, что в донной растительности доля чистой продукции фотосинтеза от валовой значительно выше, чем у других первопродуцентов рифа, таких, как кораллы или сообщества перифитона. Соотношение Р/М в ассоциациях водных растений выражается чаще всего величинами 2-4 (см. табл. 8.7), тогда как в донных биотопах с доминированием кораллов оно близко к 1,0—1,5. Важная особенность донных растительных ассоциаций рифа состоит в том, что они представлен в основном мелкоталломными или нитчатыми их формами, а также травами, которые охотно поедаются многими рыбами и беспозвоночными (Ott, Maurer, 1977). Столь же доступны для прямого выедания пышные эпифитные обрастания талломов крупных водорослей и крупнолистных морских трав, а также колонии большинства кораллин. Прямое использование потребителями значительной части первичной продукции, производимой донной флорой рифа, укорачивает пищевую цепь и на много повышает эффективность использования энергии экосистемой рифа (Randall, 1967; Bakus, 1969). Донные растения рифа служат прямым источником питания примерно четверти всех видов рыб, обитающих на рифах (Earle, 1972). Согласно Ренделлу (1967), у 59 видов рифовых рыб М. Антильских островов из 212 растительная пища составляет большую долю рациона.