Среди перечисленных таксонов известковых водорослей наиболее активными продуцентами карбонатного материала являются Halimeda и Corallinacea (Goreau, 1963; MaxweU, et al., 1964; Chave et al., 1972; Littler, Doty, 1975). Содержание извести в процентах от сухого веса достигает у халимеды 90-97%, тогда как у других известковьи зеленьи водорослей того же семейства Udoteaceae, таких, как Udotea, Penicillus, Rhipo-cephalus, оно близко к 40-60% (Bohm, et al., 1978). На большнстве рифов за счет халимеды образуется от 10 до 50% известкового материала рьильи осадков. Род Halimeda представлен на рифах Карибского бассейна преимущественно видами Н. opunta, Н. discoidea, Н. incrassata. В зоне Большого барьерного рифа Австралии преобладают Н. tuna, Н. opunta, Н. copiosa, Н. minima (Drew, Abel, 1983).
Кораллины из семейства Corallinaceae представлены на рифах несколькими родами. Пожалуй, наиболее важную роль в создании рифовых построек играют представители кораллин, образующие массивные коркообразные, иногда слабо ветвящиеся колонии, которые растут на наветренном крае рифа и на флете в участках, подверженных наибольшему волнобою. Оптимальными условиями для их роста является интенсивный водообмен и сильный свет. В составе отлагаемого ими в своих клеточных стенках кальцита содержится до 25% карбоната магния (Borowitzka, 1983). Они растут на флете, на гольх скальных поверхностях, в трещинах и выемках, а также на колониях отмершех кораллов, цементируя их. Ск орость их линейного роста достаточно велика и достигает 0,7-2 см в год (Agegian, 1981). После отмирания клеток кораллин в них начинается кристаллизация карбоната кальция с примесью магния, в результате чего отмершее участки корок кораллин подвергаются литификации, наращивая скальную основу флета (Alexandersson, 1974). Таким образом, определяющая функция корковых кораллин состоит в консолидации крупных арматурных блоков известняка и в постройке скальной основы рифа (Stoddart, 1969; Littler, 1972). Представители других родов корралин имеют вид пучков, отростков, имирегнированных кальцитом, или веточек. Водоросли Amphiroa, поселяясь на мертвых кораллах, также часто выполняют функцию цементирования. Главная же функция ветвистых кораллин состоит в продукции рыхлого карбонатного материала рифа. Продукция карбоната кальция кораллинами оценивается в 20-40% от его суммарной продукции всем сообществом рифа. Именно таково обычное для большинства рифов относительное содержание в рыхлых осадках кальцита, который образуют кораллины. Остальная масса карбонатного материала представлена арагонитной его формой, которая продуцируется кораллами и зелеными известковыми водорослями.
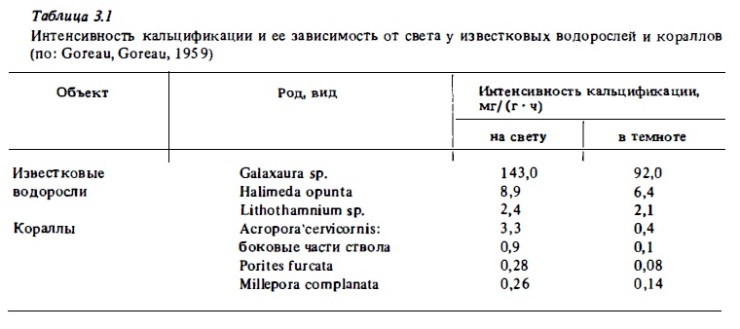
Механизм кальцификации у известковых водорослей изучался преимущественно на примерах халимеда и некоторых кораллин (Goreau, 1963; Bohm, 1978; Borowitzka, 1977; 1979). Согласно последним авторам, образование кристаллов арагонита у халимеды происходит в межклеточных промежутках наружной паренхимы таллома. Из этих промежутков, в которые из внешней среда попадают ионы гидрокарбоната кальция, происходит откачка СО2 за счет фотосинтетического ее потребления. Это и приводит к росту рН и к образованию в межклеточных промежутках СаСО3 в виде игольчатых кристаллов арагонита. Существенная особенность процесса кальцификации у халимеды состоит в ero независимости от света. На свету и в темноте этот процесс идет у нее практически с одинаковой скоростью (см. табл. 3.1). Согласно мнению Горо, именно данная особенность физиологии роста халимеды объясняет обилие разных ее видов в глубоких зонах рифа на глубинах около 60 м, где ввиду снижения освещенности резко убывает число видов кораллов и кораллин, у которых кальцификация зависит от света.
Кораллины отлагают кристаллы кальцита с большой примесью углекислого магния в своей клеточной стенке. При этом образующиеся кристаллы сразу же обрастают органическим материалом нарастающей клеточной стенки, будучи ориентированными в ней определенным образом. Это придает механическую прочность клеткам и тканям кораллин, у которых каждая клетка окружена как броней прочным известковым чехлом. Не случайно кораллины выдерживают волнобой даже там, где разрушаются массивные формы кораллов и образуют на участках края фпе ra в зонах наибольшего воздействия волн водорослевые валы, о которых упоминалось выше. Процесс кальцификации у кораллин ускоряется на свету (Borowitzka, 1979), хотя и в меньшей степени, чем у кораллов. Соотношение скорости кальцификации на свету и в темноте у кораллин, по данным Горо, составляет около 1,5 (см. табл. 3 .1), тогда как у кораллов — около 9. И все же определенная связь кальцификации с фотосинтезом прослеживается и у кораллин. Так, кальцификация у них ингибируется в присутствии химического ингибитора фотосинтеза (DCMU) так же, как и у кораллов. Возможно, за счет фотосинтеза у них происходит поставка АТФ, необходимой для активного транспорта ионов Са2+ в зону кальцификации (Barnes, Taylor, 1973). Установлено также, что в процессе образования кристаллов кальцита частично участвует углекислота, выделяемая клетками водоросли в процессе дахация (Borowitzka, 1983). Скорость кальцификации у известковых водорослей измерят путем оценки весового прироста извести, а также с помощью радиоизотопов Са4 5 и С1 4 (Borowitzka, 1977). Как уже отмечалось выше, применение Са4 5 дает завышенные результаты. Более надежные величины можно получить с помощью См (Borowitzka, Larkum, 1976), а также измеряя прямое потребление растениями ионов кальция из морской воды (Bohrn, Goreau, 1973).